|
|
|
 |
Гаркави Л.Х., Квакина Е.Б., Уколова М.А.
Адаптационные реакции и резистентность организма.
УДК 612.017
Печатается по решению бюро отделения биологии Северо-Кавказского научного центра высшей школы
Рецензенты:
доктор медицинских наук К. П. Балицкий
доктор медицинских наук М. Ф. Поляничко
Редактор Т. В. Кравченко
Ростов н/Д: Издательство Ростовского университета, 1990.-224 с.
ISBN 5- 7507- 0215- 4
В настоящее время в различных областях медицины
и биологии получила
ши-рокое распространение теория стресса Г. Селье.
Развитие стресса — реакции на разные по
качеству сильные раздражители —
лежит в основе многих
патологических процессов.
Поэтому изучение стресса вооружило медицину теорией,
позволяющей бо-лее глубоко проникать в механизмы болезней
и обосновать некоторые виды лечения.
Но, вместе с тем, появилась
тенденция распространять понятие
«стресс» на са-мые
разнообразные реакции,
вызываемые раздражителями любой силы, а не только
сильными. Это объясняется, по-видимому, тем, что
реакция стресс была единственной
известной неспецифической
адаптационной реакцией.
организма.
Однако сам характер
изменений в организме при стрессе
говорит о том, что эта реакция может быть приспособительной только по
отношению к сильным раздражителям. При
стрессе наряду с
элементами защиты имеются
элементы повре-ждения.
Приспосабливаться ценой повреждения целесообразно лишь
по отношению к сильному
раздражителю, который может
угрожать жизни. Такой путь приспособле-ния по отношению к
раздражителям обычной (слабой,
средней) силы биологически
нецелесообразен и потому не мог
закрепиться в процессе эволюции.
В данной книге говорится о найденных нами неизвестных
ранее общих неспеци-фических
адаптационных реакциях: «реакции
тренировки», развивающейся в ответ на действие разных по
качеству слабых раздражителей (Гаркави Л. X., 1969; Квакина Е. Б..,
Уколова М. А., 1969) и «реакции активации»— на
действие разных по качеству раздражи-телей средней силы (Гаркави Л. X.,
1968, 1969). Описаны комплексы изменений в
нейро-эндокринной системе, защитных системах организма и
некоторых звеньях метаболизма, характерные для каждой реакции
и отличающие их как друг от друга, так и от реакции стресс.
Подчеркивается закономерность,
объединяющая качественно
различные адаптационные реакции,
а именно, зависимость их от количества
(силы) действую-щих раздражителей.
Это количественно-качественный принцип является
методологи-ческой основой развиваемой теории
адаптационной
деятельности организма.
Из-лагаются данные, полученные
в процессе изучения найденной
3
закономерности, свидетельствующие о том, что основные адаптационные
реакции (трени-ровка, спокойная и повышенная активация, стресс) по мере
увеличения (или уменьшения) величины действующего фактора на
определенный коэффициент периодически повторя-ются на разных уровнях
реактивности организма. При этом на каждом уровне они сохра-няют свои
основные признаки (есть и отличия), и развитие каждой реакции зависит
от от-носительной, по отношению к данному уровню, величины действующего
фактора. Уро-вень же реактивности, на котором развиваются реакции,
зависит от абсолютной величины раздражителя (Гаркави Л. X., Квакина Е.
Б., 1975—1987). В монографии освещается во-прос о природе и
особенностях повышения сопротивляемости, резистентности организма при
развитии реакции тренировки и активации на разных уровнях реактивности
(в ответ на разные по абсолютной величине воздействия) по отношению к
повреждающим факто-рам внешней и внутренней среды. Приводятся простые
показатели реакций, используемые для их контроля и вызова.
Резистентность организма во многом определяет заболеваемость, тяжесть
течения болезни, эффективность лечения. В монографии показано, что,
вызывая целенаправленно нужную адаптационную реакцию организма на
определенном уровне реактивности, мож-но подойти к управлению
резистентностью организма. Возможность управления рези-стентностью с
использованием обратной связи с организмом (пользуясь разработанными
показателями) или программированных режимов, основанных на
закономерностях разви-тия реакций, открывает перспективу для применения
найденных адаптационных реакций в различных областях медицины.
Целью монографии является ознакомление широкого круга специалистов с
теорией адаптационных реакций организма и практическими рекомендациями,
позволяющими ис-пользовать реакции тренировки и активации разных
уровней реактивности для повыше-ния резистентности при лечении
различных заболеваний, активной профилактики и оздо-ровления, борьбы с
действием различных повреждающих факторов и чрезмерных нагру-зок,
вызывающих стресс, в том числе психоэмоциональных, а также для
преодоления ста-рения.
4
АДАПТАЦИОННЫЕ РЕАКЦИИ ОРГАНИЗМА
В течение всей жизни организму приходится приспосабливаться к
непрерывно ме-няющимся условиям внешней и внутренней среды. С этой
точки зрения, вся жизнь - по-стоянное приспособление.
Жизнь «на всех ступенях ее развития» И. М.
Сеченов (1863) определял «как при-способление ... к
условиям существования». Приспосабливаемость, подчеркивал Н. Е.
Введенский (1899), «составляет свойство, проникающее,
так сказать, все явления, совер-шающиеся в живом организме...»
«Постоянство внутренней среды есть условие свободной жизни
организма», по оп-ределению Клода Бернара (1878).
Уолтер Кэннон (1932) ввел термин «гомеостаз», что означает
еще более широкое понятие — поддержание относительного
динамического по-стоянства всего организма. При этом был вскрыт
механизм поддержания этого постоянст-ва -
автоматическая саморегуляция —
результат совершенствования приспособитель-ной
деятельности в процессе эволюции. В настоящее время положение о
нервно-рефлекторной регуляции гомеостаза является общепринятым и вошло
в учебные руководства (Коган А. В.,
1984).
Человек живет в условиях ледяной Арктики и знойного экватора, сохраняя
в весьма узких пределах одну и ту же температуру тела, один и тот же
состав крови и многие; дру-гие «константы».
Как же приспосабливается организм к действию бесчисленных, постоянно
меняю-щихся факторов? Как сохраняет необходимое для жизни относительное
постоянство внутренней среды?
В процессе эволюции развивались различные пути и способы
приспособления. Один из важных путей приспособления - это отсутствие
реакции организма на раздражитель. Но не менее важен другой путь
— реакции на различные раздражители. Реакции — это путь
приспособления живого к вечно меняющимся условиям, ибо устойчивость
живого во мно-гом связана с его высокой лабильностью. Приспособительные
реакции организма поддер-живают относительное динамическое постоянство
внутренней среды и функционирования всех органов и систем, необходимое
для сохранения жизни, и, с этой точки зрения, можно поставить знак
равенства между понятиями «защита» и
«приспособление», «адаптация», как в тех
случаях, когда адаптационная
реакция содержит в себе
элементы повре-ждения, так и в тех случаях, когда не содержит.
Каждый из действующих на организм раздражителей характери-
5
зуется и количеством и качеством. Что же определило формирование в процессе эволю-ции адаптационных реакций организма?
Наиболее просто представить себе качественный путь приспособления, т.
е. к каж-дому воздействию, к каждому раздражителю организм
приспосабливается путем развития различных реакций: к холоду —
одних, к химическим веществам — других, к действию электрического
тока — третьих и т. д. Но признание верным только качественного
пути приспособления приводит к абсурду. Известно, например, что в
настоящее время каждый день синтезируются сотни и тысячи новых
химических веществ, которые никогда до этого не существовали. Многие из
них входят в медицину и быт в виде лекарств, красок, пласт-масс и т. п.
и так или иначе контактируют с организмом человека, а значит, являясь
био-логически активными, воздействуют на него. Можно ли сделать выводы,
что в этой си-туации в организме ежедневно вырабатываются сотни и
тысячи новых путей приспособ-ления, новых защитных реакций? Опыт
изучения эволюции живого говорит о том, что столь существенные
изменения, как новые защитные реакции, в организме возникают крайне
медленно, в течение целого ряда поколений. Трудно себе представить,
чтобы на бесчисленное множество различных раздражителей существовало
такое же множество со-вершенно различных реакций целого организма.
Невольно напрашивается мысль о суще-ствовании не столь многочисленных
однотипных стандартных ответных реакций. Тогда каждая из таких реакций
будет являться универсальной для целой серии раздражителей, разных по
качеству, но одинаковых по силе.
Дальнейшее изучение стресса показало, что эта реакция протекает
стадийно, харак-теризуется определенным комплексом изменений в
нейроэндокринной системе и оказыва-ет влияние на уровень
неспецифической резистентности организма, его воспалительный потенциал
и обмен веществ.
Через 6 ч после стрессорного воздействия развивается первая стадия
стресса — «реакция тревоги», которая длится
24—48 ч. Эту стадию характеризует уменьшение ти-муса, лейкоцитоз,
определенное соотношение форменных элементов белой крови: лимфо-пения,
анэозинофилия, нейтрофилез и развитие кровоизлияний и язв в слизистой
желу-дочно-кишечного тракта. В эндокринной системе — выброс в
кровь адреналина надпо-чечниками, стимуляция секреции АКТГ гипофиза,
приводящая к повышению секреции глюкокортикоидных гормонов коры
надпочечников. Секреция минералокортикоидных гормонов угнетена.
Угнетены также деятельность щитовидной и половых желез. В
иссле-дованиях как зарубежных, так и отечественных авторов была
показана роль различных катехоламинов (адреналина, норадренадина,
дофамина) и их метаболизма в механизме развития стресса (Levy L., 1972;
Frankenhaeuser M., 1975; Кассиль Г. Н., Матлина Э. Ш., 1973; Шаляпина
В. Г., 1976; Диль-
6
ман В. М., 1983; Еремина С. А. и др., 1984).
После реакции тревоги наступает стадия резистентности, т. е. стадия
устойчивости. Селье установил, что в этой стадии устойчивость организма
к вредным воздействиям по-вышена. Что же касается состояния желез
внутренней секреции, то такого подробного описания их деятельности, как
в стадии тревоги, не имеется. Отмечается, что происходит некоторая
нормализация деятельности желез внутренней секреции и
тимико-лимфатической системы, а иногда даже повышение функциональной
активности желез, угнетенных в первую стадию реакции.
Селье показал, что если раздражитель (стрессор) очень сильный или
повторяется, то стадия резистентности
переходит в стадию истощения. Сильное или
длительное напряжение приводит к повторному угнетению защитных систем
организма. Характер деятельности эндокринных желез очень близок к тому,
что наблюдается при реакции тре-воги: глюкокортикоиды преобладают над
минералокортикоидами, снижена
активность щитовидной железы
и половых желез, угнетена тимико-лимфатическая система,
систе-ма соединительной ткани, иммунная система. Однако, если при
реакции тревоги отмеча-ется резкое повышение секреции АКТГ и
глюкокортикоидных гормонов, то в течение ста-дии истощения секреция
глюкокортикоидов начинает снижаться и, наконец, тоже падает.
Как понять значение и последовательность отдельных фаз при
стрессе?
В чем биологический смысл первой стадии — реакции тревоги? При
встрече с сильным раздражителем основная задача — любой ценой
получить энергию в короткие сроки, чтобы обеспечить необходимые условия
для «битвы» или «бегства». Быстрый вы-брос
энергии мобилизуется адреналином и глюкокортикоидами даже невыгодным
путем за счет распада жиров, углеводов и белков (прежде
всего, лимфоидной ткани). Глюко-кортикоиды в больших количествах
угнетают тимус, лимфатические
железы, произ-водство лимфоцитов,
иммунные реакции, а
также оказывают противовоспалитель-ное
действие, т. е. подавляют деятельность защитных систем организма
(известно, что воспалительная реакция в процессе эволюции
возникла как защитная реакция).
Мине-ралокортикоидные гормоны, которые оказывают противоположное
влияние на течение воспалительного процесса, напротив, угнетены. Эти
изменения биологически целесооб-разны, так как защитный ответ,
адекватный большой силе раздражителя (например, вос-палительная
реакция), мог бы привести организм к гибели. Если бы не развивалась
имму-нодепрессия, то при стрессе в условиях повреждения тканей в
постстрессорный период могли бы возникнуть аутоиммунные
заболевания (Дильман В. М., 1969).
Поэтому вна-чале организму
приходится не усиливать, а ослаблять свой
ответ. Складывается пара-доксальное, на первый
7
взгляд, положение: в ответ на действие сильного раздражителя активность
основных за-щитных систем не нарастает, а напротив, подавляется. Таким
образом, изменения в эн-докринной системе в реакцию тревоги
представляют собой, по сути дела, не «призыв к оружию», а
«призыв к разоружению», так как приводят к подавлению
активности защит-ных систем организма. Вместе с этим повышается
артериальное давление и свертывае-мость крови — это обеспечивает
готовность к борьбе и возможным кровотечениям при ранениях. Иные же
функции, например пищеварения, так же как и активность других же-лез
внутренней секреции (половых, щитовидной), ненужные для
непосредственной борь-бы с сильным раздражителем,— подавлены. Все
эти приспособительные изменения, про-исходящие в первую стадию стресса,
могут вызвать тяжелые последствия в организме, особенно в условиях
гипокинезии и гиподинамии, когда изменения, присущие стрессу, не
реализуются на мышечную работу (Эверли, Розенфельд, 1985). Поэтому
необходимо пом-нить, что реакция тревоги — это типичный пример
того случая, когда защита достигается ценой повреждения.
Но как представить себе, почему вслед за реакцией тревоги, т. е. на
фоне угнетения защитных систем организма, формируется без каких-либо
дополнительных воздействий стадия резистентности, т. е. происходит
нормализация или даже повышение устойчиво-сти? Известно, что в ЦНС под
влиянием сильных раздражителей развивается резкое воз-буждение, которое
затем сменяется запредельным торможением — «крайней мерой
защи-ты» по И. П. Павлову. При запредельном торможении
чувствительность центральных нервных аппаратов снижается. Создается
впечатление, что именно благодаря этому па-дающие на организм другие
сильные воздействия уже воспринимаются не как сильные, и тем самым
устойчивость организма повышается. Мы связываем переход стадии тревоги
в стадию резистентности с развитием в ЦНС запредельного торможения.
Стадия истощения еще в большей степени, чем стадия тревоги, является
примером такого состояния, когда сохранение жизни достигается ценой
повреждения. В наиболее тяжелых случаях эта стадия может привести к
гибели.
Какое свойство раздражителей может создать нечто общее в ответной
реакции на разные по качеству раздражители, сформировать основу для
стандартного приспособи-тельного ответа? Качество не может явиться
такой основой, так как каждому раздражите-лю присуще свое качество.
Общее, что характеризует действие самых различных
раздра-жителей,— это количество, определяемое в отношении живого
как степень биологической активности. Раздражителям, разным по
качеству, может быть присуща одна и та же сте-пень биологической
активности (одно и то же количество), а раздражителям, одинаковым по
качеству,— разная степень биологической активности (разное
количество).
8
Разумеется, представление о чисто количественном пути приспособления
без учета качественных особенностей раздражителей также противоречит
фактам. Однако количе-ство, мера может явиться основой общности реакции
организма на действие разных по качеству раздражителей, основой для
развития в процессе эволюции биологически
целе-сообразных комплексных,
стандартных ответных реакций организма.
По нашему мнению, в этой основе лежит количественно-качественный
принцип: в ответ на действие раздражителей, различных по количеству,
т. е. по степени своей биоло-гической активности, развиваются
различные по качеству стандартные
адаптационные реакции организма. Иными словами, мы считаем, что
развившиеся в процессе эволюции общие приспособительные реакции
организма являются неспецифическими, а специфика, качество каждого
раздражителя накладывается на общий неспецифический фон.
Общие адаптационные реакции являются реакциями всего организма,
включаю-щими в себя все его системы и уровни. Это создавшиеся в
длительном процессе эволюции комплексные защитные реакции. С целью их
изучения можно исследовать в отдельности изменения в какой-то одной
системе или на каком-то одном, например молекулярном, уровне. Но нельзя
забывать, что это только часть изменений в общей комплексной реак-ции
организма.
Чем характеризуются общие
неспецифические адаптационные реакции орга-низма и как они
формируются? Эти комплексные реакции характеризуются, прежде всего,
автоматизмом. Как осуществляется такая автоматическая саморегуляция?
Важнейшая роль в приспособлении
принадлежит ЦНС —
основной регулирующей системе ор-ганизма. Кора
головного мозга с системой анализаторов принимает информацию от
внешнего мира, подкорковые образования головного мозга — от
внутренней среды. Ав-томатическое регулирование
постоянства внутренней среды
осуществляется, глав-ным образом, гипоталамической областью
мозга, являющейся центром интеграции веге-тативного отдела
нервной системы и эндокринной системы — основных исполнительных
звеньев, реализующих влияние ЦНС на внутреннюю среду организма (Алешин
Б. В., 1971, 1976, Тонких А. В., 1965). В гипоталамусе сочетаются
нервный и гуморальный путь
автоматической регуляции. Н.
И. Гращенков (1965) образно сравнил
гипота-ламус с радарной установкой, включенной в систему
саморегулирования и автоматиза-ции нейрогуморально
гормональных процессов, противостоящих
динамически изме-няющимся факторам не только внутренней, но и
внешней среды О роли гипоталамиче-ских образований в формировании
стресса свидетельствуют и данные о ранних нейрохи-мических
и физиологических изменениях в гипоталамо-гипофизарной системе при
стрес-сорных воздействиях (Хайдарлиу С. X., 1984; Фурдуй Ф. И., 1986)
9
Наличие теснейшей анатомо-физиологической связи между гипоталамусом и
рети-кулярной формацией, играющей важную роль в осуществлении
генерализованных неспе-цифических реакций, также говорит о значении
этих отделов мозга в формировании не-специфических реакций организма.
Общая неспецифическая реакция организма
на сильные раздражители — стресс (реакция напряжения)
Первая общая неспецифическая адаптационная реакция была открыта
канадским ученым Г. Селье. Им было показано, что в ответ на действие
разных по качеству, но силь-ных раздражителей в организме стандартно
развивается один и тот же комплекс измене-ний, характеризующий эту
реакцию, названную общим адаптационным синдромом (ОАС), или реакцией
напряжения — реакцией стресс (Селье Г., 1936-1972).
Селье, еще будучи студентом медицинского института, обратил внимание на
то, что у многих совершенно различных болезней признаки очень сходны. В
дальнейшем, изучая на животных действие экстрактов из плаценты и
яичников, Селье через 6 ч после введения обнаружил у всех животных
увеличение коркового слоя надпочечников (с ис-чезновением
секреторных гранул из корковых клеток), инволюцию тимико-лимфатического
аппарата и кровоточащие язвы желудка и двенадцатиперстной кишки.
Вначале Селье предположил, что им открыт новый гормон, который
оказывает такое дей-ствие. Однако по мере того, как Селье
«очищал» вытяжку, стремясь получить гормон в чистом
виде,— все изменения постепенно уменьшались. Нужно отдать должное
Селье, который в такой трудный для исследователя момент сумел отойти от
привычного пред-ставления о главной роли качества воздействия и подумал
о том, что в неочищенном виде вытяжка была просто более сильным
раздражителем по сравнению с очищенной, и реак-ция на нее — это
реакция на сильный раздражитель. Если это так, то любой другой силь-ный
раздражитель должен был дать аналогичный эффект. Селье ввел животным
формалин и получил те же изменения. Именно в связи с этим он вспомнил
свои студенческие впе-чатления о сходстве многих признаков при разных
болезнях, возникающих, как известно, в результате действия сильных
агентов. Как говорил И. П. Павлов (1927), «патологическое
состояние есть встреча организма с каким-нибудь чрезвычайным условием,
вернее — с необычным размером ежедневных условий».
Все многочисленные исследования действия на метаболизм различных
экстре-мальных факторов говорят о том, что эти воздействия вызывают
большие энергетические траты и преобладание процессов катаболизма над
процессами анаболизма. Поскольку стадия рези-
10
стентности может наступать только после реакции тревоги, которая
протекает с большими энергетическими тратами, элементами повреждения и
угнетения защитных систем орга-низма, то повышение резистентности при
стрессе достигается, по словам самого Г. Селье,
«дорогой
ценой».
Таким образом, реакция на любой по качеству, но сильный раздражитель
содержит и элементы защиты, и элементы повреждения. Такая реакция на
сильный раздражитель биологически целесообразна, так как сильный
раздражитель может угрожать жизни, за-щищая которую организм должен
идти на любые траты и жертвы.
Вначале Селье описал острый стресс, т. е. реакцию, развивающуюся в
ответ на дей-ствие сильного раздражителя — однократное или
эпизодическое. Это может быть сильное переохлаждение, перегревание,
хирургическая операция и т. д.
Но вернемся к тому, с чего начал Селье-студент: к болезням. При
хронических за-болеваниях действие сильного, повреждающего раздражителя
длится очень долго. Если при острых инфекционных заболеваниях действие
болезнетворного агента длится дни и недели, то при хронических —
это месяцы, иногда годы, а при опухолевой болезни — все
время, пока опухоль находится в
организме. При острых инфекциях еще удается четко отметить
«реакцию тревоги». При хронических заболеваниях ее замечает
не всегда, а при развитии опухолей большей частью больной обращается к
врачу, когда в той или иной степени развивается стадия истощения. Все
это стресс, но особый — растянутый во вре-мени, хронический
стресс, причем растянут он в основном за счет длительной стадии
ис-тощения. Наши исследования показали, что стадия истощения
хронического стресса про-текает волнообразно. Например, признаки
истощения секреции глюкокортикоидов то на-растают, то
уменьшаются. Возможно, это связано с тем, что
возбудимость центральных нервных структур волнообразно
колеблется от резкого возбуждения до запредельного торможения, включая
и «фазовые состояния», когда чувствительность
извращается.
Селье было четко показано наличие общей неспецифической реакции стресс
в от-вет на разные по качеству, но сильные раздражители. Спустя почти
40 лет после открытий ОАС, Селье (1973) пишет, что медицине
понадобилось весьма длительное время, чтобы принять существование
однотипной реакции на действие различных, практически любых
раздражителей, что на первый взгляд казалось
нелогичным.
Однако нужно отдать должное Селье, которому не только удалось убедить
медиков в существовании такой реакции, но и создать одну из самых
популярных теорий в меди-цине. Учение о стрессе получило широкое
распространение во всем мире. Появилось ко-лоссальное количество
работ о влиянии стрессорных воздействий на
11
различные органы и системы организма. По мере накопления громадного
числа факторов представление Селье о системе «гипофиз —
кора надпочечников» как основной системе, повышение активности
которой приводит к развитию стресса, несколько изменилось. Се-лье
пришел к выводу, что усиление секреции глюкокортикоидов во время
реакции тревоги не обязательно. Реакция тревоги может сформироваться и
при «почти физиологических количествах» гормонов,
требующихся лишь для создания определенного гормонального соотношения.
Говоря о роли АКТГ, Селье подчеркивает, что важна не только абсолютная
величина секреции АКТГ, но и соотношение с СТГ. Однако большинство
исследователей говорит о повышении секреции АКТГ и глюкокортикоидов при
стрессе (Selye H., 1950, 1956, 1960, 1961; Selye H., Horava A., 1953;
Горизонтов П. Д., Протасова Т. Н., 1968; Ере-мина С. А., 1973; Митюшов
М. И., 1976; Филаретов А. А., 1979).
Кроме того, Селье пришел к выводу об участии и других систем в
формировании реакции стресс. Факты, показывающие, в частности, что
типичный трехстадийный стресс может быть воспроизведен на
адреналэктомированных крысах, правда, в несколько видо-измененной форме
(непродолжительная стадия резистентности, быстро наступающая ста-дия
истощения), привели Селье к мысли об участии щитовидной железы в
развитии реак-ции стресс и о том, что не все виды неспецифической
резистентности опосредуются через единый гормональный механизм. В
качестве таких новых возможных механизмов он на-зывает систему
«гипофиз — щитовидная железа» (Selye Н., 1976).
На основании исследований гормональной активности при реакции избегания
и иммобилизационном стрессе у обезьян Мейсон сделал вывод об участии в
развитии стрес-са щитовидной, поджелудочной и половых желез, причем
особое внимание обратил на то, что уровень катаболических гормонов
повышается, а анаболических — снижается. Так, помимо
глюкокортикоидов, адреналина и норадре-налина, повышалась секреция
гормона роста и гормонов щитовидной железы, а секреция половых гормонов
(эстрогенов и андро-генов) снижалась (Mason J. W., 1975). Данные по
вопросу об изменении активности щито-видной железы противоречивы.
Большинство исследователей отмечает снижение функ-ции щитовидной железы
при стрессе (Туракулов Л. X., 1972; Дильман В. М.,
1986; и др.).
Однако, несмотря на развитие и изменение первоначальных взглядов Селье,
оста-ется незыблемым положение о наличии при всех видах стресса реакции
тревоги с лимфо-пенией, эозинопенией, лейкоцитозом, инволюцией тимуса,
наличием язв в желудочно-кишечном тракте (именно эти признаки позволили
Селье в 1936 г. обнаружить «общий адаптационный синдром»).
Деление стресса на «хороший» (эустресс) и
«плохой» (ди-стресс) не меняет
положения, так как
12
не описаны конкретные отличия этих видов стресса по комплексу изменений
в нейроэн-докринной или в других системах организма (Селье Г., 1982).
По-видимому, эустресс - это либо более мягкий стресс с менее
выраженными элементами повреждения, либо другая более физиологичная,
чем стресс, и ранее неизвестная адаптационная неспецифическая реакция
организма, которую принимают за стресс
Для отечественных ученых, воспитанных на идеях И. П. Павлова, Л. А.
Орбели, Н. Е. Введенского, А. Д. Сперанского, П. К. Анохина, показалось
бы странным, что в общей приспособительной реакции организма участвует
только одна, хотя и важная система «ги-пофиз — кора
надпочечников». Мы считаем, что в осуществлении общей
не-специфической приспособительной реакции принимают участие все
системы организма, и в первую очередь — регулирующие, как
показано Г. И. Косицким и В. М. Смирновым на примере стресса (1970)
Селье в последние годы исследовал роль ЦНС в механизме разви-тия
стресса и пришел к выводу, что именно в ЦНС формируется начальный этап
стресса, дальнейшее развитие которого реализуется через подчиненные
системы организма нерв-ным, и гуморальным путем, г частности, через
эндокринную систему.
Учение о стрессе получило такое широкое распространение по тому, что
медицин-ская наука очень нуждается в теории, позволяю щей объединить в
единое целое множест-во отдельных фактов. «В руках
ученых,— писал И.-В. Гете,— часто
бывают части, но очень редко — священная
связующая нить». Селье нашел эту нить, и одной из его важ-нейших
заслуг является утверждение теории в медицине. Со свойственной ему
эмоцио-нальностью Селье писал: «Больше всего я хочу подчеркнуть
значение теории. Это осо-бенно важно сделать поскольку в наше время
испытывают просто ужас перед абстракт-ным мышлением в медицине».
Однако «ухватившись» за теорию стресса, многие ученые тут
же перестали прида-вать значение тому, что стресс — это реакция
на стрессор, чрезвычайный раздражитель, а не на любой раздражитель
вообще, что Селье пришел к мысли о стрессе отчасти и потому, что
заметил общие признаки при самых различных заболеваниях, т. е.
чрезвычайных об-стоятельствах для организма. Кроме того, многие
исследователи даже не изучают, разви-вается ли после воздействия
комплекс изменений, характерный для стресса, заведомо счи-тая, что
любой раздражитель вызывает стресс. Если же при этом обнаруживается
хоть ка-кое-либо изменение деятельности «системы гипофиз —
кора надпочечников», то исчеза-ют и последние сомнения (если они
были), стресс это или не стресс. Между тем система «гипофиз -кора
надпочечников» является одним из важных звеньев регуляции
организма. Неужели эта система работает только в чрезвычайных условиях,
а в обычных физиологи-ческих полностью бездействует? Ясно, что
регулирующие системы работают всегда, но по-разному, в зависимости от
условий.
13
Селье в большинстве своих работ говорит о том, что стресс — это
реакция на силь-ный раздражитель, но вместе с тем четкого разделения
раздражителей по силе не прово-дит. Это приводит к путанице, к
представлению, что стресс — общая неспецифическая адаптационная
реакция на любой раздражитель.
Трудно представить себе, чтобы для приспособления не к чрезвычайным, а
слабым и средней силы раздражителям было бы биологически целесообразно
подавление естест-венного защитного ответа, к тому же ценой повреждения.
О возможности существования других (кроме стресса) неспецифических адаптаци-онных реакций организма
П. Д. Горизонтов и Т. Н. Протасова (1968) полагают, что Селье
необоснованно стремится безгранично расширить понятие стресса, включая
в него все неспецифические реакции организма. Действительно, нельзя
забывать, что реакция стресс — это общая
не-специфическая адаптационная
реакция на
сильный раздражитель. И. А. Аршав-ский считает
стресс, описанный Селье, «патологическим стрессом» и
говорит о необхо-димости выделения «физиологического
стресса» (1976, 1982, 1986). Однако он четко не связывает
развитие «физиологического стресса» с определенной
величиной действующего фактора, хотя и отмечает, что она не должна быть
чрезмерной. Наличие множества
различных приспособительных реакций организма в
ответ на действие многочисленных разных по качеству раздражителей
представляется невозможным. В такой же степени ма-ловероятным
представляется наличие лишь одной реакции
на действие раздражителей разной силы. Особый интерес в связи с этим
имеют исследования Н. В. Лазарева (1962, 1963) и его
последователей по изучению состояния неспецифически повышенной
сопротивляемости организма (СНПС).
Это состояние развивается под влиянием группы ве-ществ
(элеутерококка колючего, женьшеня, дибазола и т. д.), объединенных
Лазаревым под названием «адапто-гены». Сравнение ОАС
и СНПС показало, что при развитии СНПС отсутствуют признаки полома,
заметные изменения (по сравнению с нормой) в системе
«гипофиз — надпочечники» и в щитовидной железе,
нет существенных измене-ний тимуса, отмечается лишь активация половых
желез и гонадотропной функции гипо-физа (Кириллов О. А., 1964; Розин М.
А., 1966). Лазарев считает это состояние качествен-но отличным от ОАС,
полагая, что СНПС достигается физиологическим путем. Много-численные
исследования показали, что адаптогены в применяемых дозах повышают
не-специфическую ре-зистентность и защищают организм от действия
стрессоров (Брех-ман И. И., 1957, 1968, 1987; Кириллов О. А.,
1964; Розин М. А., 1966; Дардымов И. В., 1976; и др.)- Большой заслугой
Н. В. Лазарева
14
является то, что он первый сказал о возможности существования
качественно иного, чем стресс, состояния организма, также приводящего к
повышению неспецифической рези-стентности, но достигаемого
физиологическим путем.
Особенно удачным был выбор Лазаревым и его последователями веществ,
являю-щихся биостимуляторами широкого спектра действия с отсутствием
выраженного специ-фического и, особенно, токсического влияния. Однако
нельзя согласиться с тем, что такое неспеци-фическое
состояние, как СНПС,
может быть достигнуто только действием
определенной группы в этом смысле специфических веществ —
адаптогенов. Те же адап-тогены в зависимости от дозы могут вызывать и
СНПС, и стресс. Так, применение боль-ших доз дибазола и женьшеня
приводило к изменениям в эндокринной системе, характерным не для СНПС,
а для стресса (Розин М. А., 1966; Петков В., 1974; Дардымов И. В.,
1976;и др.)- И напротив, как будет показано, самые разнообразные
раздражители, не отно-сящиеся к адаптогенам, способны
вызывать физиологическим путем
реакции орга-низма, отличные от
стресса, и повышать его
неспецифическую резистентность.
Таким образом, мы опять возвращаемся к уже высказанному нами ранее
количест-венно-качественному принципу приспособительной деятельности
организма: развитию качественно различных реакций организма в
зависимости от меры, количества раздражи-теля. Хочется еще раз
подчеркнуть значение количества, меры как того общего, что
ха-рактеризует действие самых различных по качеству раздражителей.
Именно поэтому ко-личество (мера) могло явиться основой для
формирования нескольких стандартных, а не бесчисленного множества
разного типа приспособительных реакций организма. Чтобы убедиться в
правильности количественно-качественного принципа приспособления,
необ-ходимо было исследовать действие на организм раздражителей
различной силы.
Наши исследования показали, что в зависимости от силы (дозы)
воздействия, в организме могут развиваться минимум 3 адаптационные
реакции:
1) реакция на слабые воздействия — реакция
тренировки (Гаркави Л. X., 1969; Квакина Е. В., Уколова М. А.,
1969);
2) реакция на воздействия средней силы, промежуточной
между слабыми и сильными — реакция активации (Гаркави Л.
X., 1968, 1969), подразделенная на спокойную и повы-шенную;
3) реакция на сильные,
чрезвычайные воздействия — стресс Селье.
Для доказательства того, что кроме известной общей неспецифической
адаптаци-онной реакции на разные по качеству, но сильные раздражители
— стресса, существуют другие общие неспецифические
адаптационные реакции, нужно
было, во-первых, вызвать
15
эти реакции, варьируя силу (дозу) раздражителей, и, во-вторых,
показать, что найденные реакции отличаются и от стресса, и друг от
друга, т. е. что они действительно разные.
Какие раздражители целесообразно было использовать?
Поскольку эти реакции комплексные, мы считали, что их легче всего
вызывать, из-меняя функциональное состояние ЦНС — системы,
регулирующей деятельность целого организма. Так как это реакции
автоматические, поддерживающие постоянство внутрен-ней среды организма,
регуляция которого осуществляется преимущественно гипоталами-ческой
областью головного мозга, то мы отдавали предпочтение воздействию на
гипота-ламус. Учитывая большую роль ретикулярной формации в
осуществлении генерализован-ных неспецифических реакций, мы
использовали некоторые средства, являющиеся ее раз-дражителями. Кроме
того, применялись биологически активные малотоксичные препара-ты
широкого спектра действия (различные биостимуляторы). Нами были
применены сле-дующие воздействия:
1. Электрическое раздражение (различной
силы) гипоталамуса через хрониче-ски вживленные электроды,
поскольку известно, что локальное электрическое раздраже-ние
гипоталамуса вызывает комплексные поведенческие реакции (Коган А.
Б., 1952).
2. Постоянные и переменные низкочастотные магнитные поля
различных парамет-ров — воздействия, главным образом, на
голову— как способ бесконтактного изменения функционального
состояния ЦНС, прежде всего гипоталамуса, обладающего высокой
чувствительностью к магнитному полю, т. е. как способ влияния на
функциональную ак-тивность гипоталамо-гипофизарной системы с включением
неспецифических механизмов адаптации (Холодов Ю. А., 1966,
1975; Becker R., 1969; Persinger M., 1976; Васильев Н. В., 1975;
Удинцев Н. А., 1976; Нахильницкая 3. Н., 1982, и др.).
3. Нейротропные средства: адреналин и адреномиметики,
холи-номиметики, ан-тидепрессанты
(мелипрамин), этимизол, центро-феноксин.
4 Биостимуляторы (в разных дозах) растительного и животного
происхождения (элеутерококк колючий, золотой корень, пантокрин, корень
левзеи, мумие, прополис, женьшень, сборы трав).
5 Углекислый газ (изменение концентрации во вдыхаемом
воздухе) как раздра-житель дыхательного центра,
ретикулярной формации и интерорецепторов, играющих
важную роль в регуляции внутренней среды организма (Черниговский
В. Н., 1960).
6. Иммуномодуляторы (левамизол, тималин).
7. Антиоксиданты (ионол, витамин Е).
8. Янтарная кислота и ее соли.
9. Дозированные физические нагрузки (плавание, бег,
ходьба). С помощью этих воздействий, варьируя их силу (дозу), мы
вызывали
16
развитие различных адаптационных реакций и исследовали в одни и те же
сроки функ-циональное состояние ЦНС, эндокринных желез,
тимико-лимфатической системы и
некоторые показатели метаболизма. Прежде всего мы изучали
те показатели, на основа-ние закономерных изменений которых Селье
удалось обнаружить стресс. Исследовали также уровень и характер
неспецифической резистентности организма при развитии раз-ных
адаптационных реакций по степени его защиты от действия различных
повреждаю-щих факторов внешней и внутренней среды. Полученный
фактический материал обрабо-тан с помощью параметрических и
непараметрических методов вариационной статистики. Только при наличии
статистической значимости отличий говорится об уменьшении или
увеличении изучаемого признака.
В результате этих исследований выяснилось, что кроме общей
неспецифической реакции на сильные раздражители — стресса,
существуют также общая неспецифическая адаптационная реакция на слабые
раздражители, названная нами "реакцией тренировки", и общая
неспецифическая адаптационная реакция на раздражителя средней силы,
назван-ная Л. Х.Гаркави «реакцией активации».
Реакция тренировки — общая неспецифическая адаптационная реакция на слабые раздражители
Как приспосабливается организм к слабым раздражителям? Существует ли
общая неспецифическая адаптационная реакция на слабые раздражители
независимо от их каче-ства подобно тому, как существует реакция стресс
на разные по качеству сильные раздра-жители?
Исследования показали, что такая реакция есть. Общая неспецифическая
адаптаци-онная реакция, развивающаяся в ответ на разные по качеству
слабые раздражители, на-звана реакцией тренировки (Гаркави Л. X., 1968,
1969; Квакина Е. В., Уколова М. А., 1969). Реакция тренировки так же,
как и стресс, характеризуется определенным комплек-сом изменений в
нейрозндокринной системе. Эта реакция протекает стадийно, и каждой ее
стадии свойствен определенный уровень неспецифической резистентности
организма.
Чем характеризуется реакция тренировки? Как она развивается?
В природе и повседневной жизни встречается громадное количество слабых
раз-дражителей, на которые организм реагирует, хотя они и слабые. Но
может ли это быть та-кая же реакция, как на сильные раздражители? Есть
ли смысл, например, в подавлении защитных систем, в защите от слабого
раздражителя ценой повреждений Ответ напраши-вается сам собой: ведь
слабые раздражения не угрожают жизни, и, может быть, было бы
целесообразно не реагировать на них вообще. Но организм должен вначале
проанализиро-вать, убе-
17
диться, что это слабый раздражитель, т. е.
«сориентироваться», и «на всякий случай»
при-вести свои защитные системы в состояние «боевой
готовности». Это первая стадия в ре-акции организма на слабое
воздействие, названная нами стадией «ориентировки». Как и
стадия тревоги стресса, она формируется через 6 ч и длится 24—48
ч после воздействия. Однако характер изменений совершенно иной. Если
для реакции тревоги характерно уменьшение тимуса, то в стадии
ориентировки тимус несколько (в 1,2—1,3 раза) увеличен по
сравнению с контролем (животные без специальных воздействий,
находящиеся в раз-личных реакциях) и находится в пределах нижней
половины зоны нормы (рис. 1). Срав-нение величины
тимуса
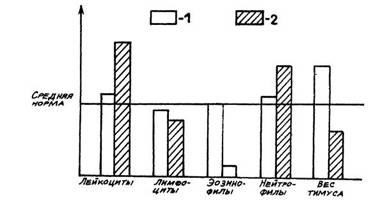
Рис. 1. Лейкоциты, лейкоцитарная формула и масса тимуса в первые стадии реакций тренировки (1) и стресса (2)
животных, находящихся в стадии ориентировки и реакции тревоги,
показало, что в пер-вую стадию реакции тренировки тимус больше в
2—3 раза, чем в первую стадию стресса. Микрокартина тимуса при
реакции тренировки соответствует классическому описанию • в
руководствах по гистологии (рис. 2). Ширина корковой части дольки в 2
раза уже мозго-вой. Гемоцитобласты, обладающие высокой степенью
пиронинофилии, располагаются преимущественно по периферии корковой
части и на границе с мозговой. В последней видны единичные тельца
Гассаля.
В лимфатических узлах крыс, находившихся в реакции тренировки,
архитектоника узла также соответствует микрокартине, приводимой в
классических руководствах (рис. 3). В селезенке герминативные центры
лимфоидных узелков состоят преимущественно из лимфобластов, больших и
средних лимфоцитов. Ширина герминативных центров превы-шает ширину
короны зрелых лимфоцитов (рис. 4).
При реакции стресс (рис. 5) корковая часть тимуса в одних слу-
18
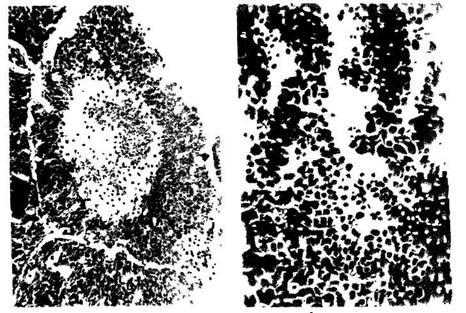
Рис 2. Тимус крыс. Реакция тренировки: отсутствие признаков как гипер-,
так и гипоплазии лимфоидной ткани. Окр по Браше в модификации А А.
Симаковой Ув. 8X7
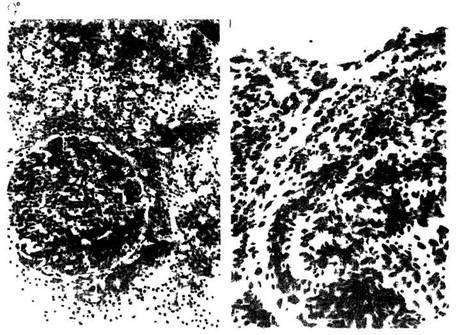
Рис 3 Лимфатические узлы крыс. Реакция тренировки, примерно равное
соотношение промежуточных синусов и мякот-ных тяжей. Окр по Браше
в модификации А А. Симаковой Ув.
8X7
чаях сужена, в других — в ней видны очаговая или тотальная
гипоплазия пиронинофиль-ных и малых лимфоцитов с выраженной
дегенерацией, увеличение числа тучных клеток в междольковой
соединительной ткани. В мозговом веществе нарастает число телец
Гассаля (Гаркави Л. X., 1968, 1969; Квакина Е. Б. 1972; Мулатова
А.К. 1983). Для лимфоуз-лов крыс в стрессе (рис. 6)
характерны гипоплазия лимфоидной ткани, истончение мя-котных тяжей,
расширение синусов, резкое ослабление пиронинофилии цитоплазмы,
усиление макрофагальной реакции, увеличение числа
тучных клеток. При стрессе в се-лезенке (рис. 7)
уменьшаются число и размеры фолликулов, во многих из них стирается
грань между центром и поясом зрелых лимфоцитов. Такая же картина
отмечена и в пери-артериальных лимфоидных муфтах.
Стадию ориентировки так же, как и реакцию тревоги, характеризуют
определенные соотношения форменных элементов белой крови (см. рис. 1).
Как отмечалось ранее, при реакции тревоги наблюдается лейкоцитоз,
анэозинофилия, лимфопения. Нами показано, что у крыс
19

Рис 4. Селезенка крыс Реакция тренировки: широкие терминативные
центры, узкие средние и наружные пояса фолликулов. Окр. по Браше
в модификации А А Симаковой
Ув 16X7
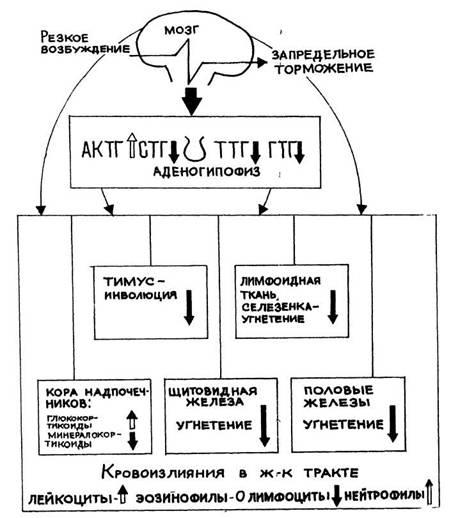
Рис. 5. Тимус крыс Реакция стресс:
резко выраженная дегенерация лимфоидных
элементов. Окр но Браше в модификации А.
Симаковой. Ув. 8х7
в стадии реакции тревоги острого стресса наблюдаются лейкоцитоз (более
20• 109), анэо-зинофилия, число лимфоцитов ниже 45%,
сегментоядерных нейтрофилов — выше нормы, число палочкоядерных
нейтрофилов и моноцитов — в норме или выше нормы. В стадии
ориентировки у крыс число лимфоцитов находится в пределах нижней
половины зоны нормы (45—60%), число сегментоядерных нейтрофилов
— в пределах верхней половины зоны нормы, число лейкоцитов,
эозинофилов, палочкоядерных нейтрофилов и моноцитов — в пределах
нормы. У людей наблюдаются следующие параметры реакции тревоги при
остром стрессе число лимфоцитов менее 20%, число лейкоцитов болee
8,0*109, анэозино-филия, число сегментоядерных нейтрофилов выше нормы,
палочкоядерных нейтрофилов и моноцитов - в пределах нормы и выше. В
стадии ориентировки у людей число лимфоци-тов находится в пределах
нижней половины зоны нормы (21 — 27%), сегментоядерных
нейтрофилов — в пределах верхней половины зоны нормы, число
лейкоцитов, эозинофи-лов, палочкоядерных
нейтрофилов и моноцитов — в
пределах нормы
20
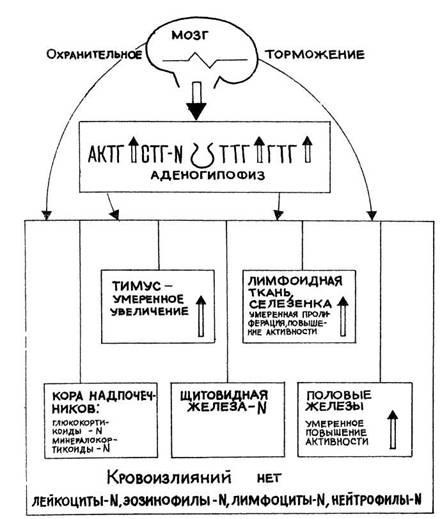
Рис. 6 Лимфатические узлы
крыс. Реакция стресс: участки дегенерации и
гипоплазия лимфоидных эле-ментов Окр по Браше в модификации
А. А. Симаковой
ув. 8X7

Рис. 7. Селезенка крыс. Реакция
стресс: увеличение числа мегакариоцитов в красной
пульпе. Oкр. по Браше в модификации А. А.
Симаковой Ув 40X7
(Гаркави Л X , 1968, 1969, Гаркави Л. X , Квакина Е Б , Уколова М А , 1974, 1975)
При оценке адаптационных возможностей организма важное значение
придается изучению энзимоцитохимических показателей форменных элементов
крови, в частности, дегидрогеназу лимфоцитов, поскольку они являются
показателями функционального со-стояния клетки и установлена коррекция
этих показателей с активностью многих систем организма (Нарциссов Р. П
, 1975, Комиссарова И.А. и др . 1978, Яновская А С , 1973;
Шепотиновский В. И , Микашинович 3 И, 1980). Исследовали
следующие ферменты: сукцинатде-гидрогеназу (СДГ, К Ф 1 3 99 1), уровень
активности которой отражает ин-тенсивность
работы цикла Кребса,
лактатдегидрогеназу (ЛДГ, К Ф1.1.1. 27),
уро-вень активности которой
характеризует интенсивность
гликолитических процес-сов,
а-глицерофосфатдегидрогеназу цитоплазматическую и митохондриальную
(?-ГФДГ цит, К Ф 1.1.1. 8, ?- ГФДГмит, К.Ф. 1.1.99. 5), уровень
активности которых
21
характеризует сопряженность процессов дыхания и гликолиза. Кроме того,
активность ? -ГФДГ в настоящее время рассматривают как показатель
активности гена иммунного отве-та (Зарецкая Ю. М., 1983), При стрессе
снижена активность СДГ и в разной степени повышена активность
ЛДГ, т. е. отношение ЛДГ/СДГ всегда увеличено; активность ?
-ГФДГцит и ? -ГФДГмит либо не изменена, либо повышена (особенно ?
-ГФДГцит); при реакции тренировки активность СДГ, ЛДГ и ? -ГФДГцит
— в пределах нижней половины зоны нормы, а ? -ГФДГмит —
несколько выше (Котляревская Е. С., Марьяновская Г. Я., Барсукова Л.
П., Коробеиникова Е. П., Мамцева Л. И., 1982).
Стадия ориентировки, как и реакция тревоги стресса (рис. 8),
характеризуется определен-ным комплексом изменений в эндокринной
системе (рис. 9). Умеренно увеличивается секреция и глюко-кортикоидных,
и минералокортикоидных гормонов коры надпочечников. Через 6 ч после
однократного слабого воздействия содержание 17-ОКС в крови лю-дей
—15,1 ± 1,1 мкг%; в то время как при реакции тревоги
стресса—28,3±1,8 мкг%; в кон-троле —9,2 ±
0,85 мкг%. Содержание 11-ОКС в крови крыс после однократного слабого
воздействия—42,7 ± 3,8 мкг%; при реакции
тревоги—69,9±5,1 мкг%; контроле без специ-ального
воздействия—31,2 ± 4,8 мкг%. Исследования у людей выявили,
что содержание кортизола в крови при реакции тренировки—385
± 81,7 нмоль/л, а при стрессе—531 ± 92,3 нмоль/л.
Морфофункциональное исследование коры надпочечников у крыс показало,
что при стрессе в стадии реакции тревоги кора надпочечников значительно
увеличена за счет пуч-ковой зоны, ответственной за секрецию
глюкокортикоидов (рис. 10) отмечается усиление вакуолизации цитоплазмы
клеток мозгового вещества. Ширина пучковой зоны 562,5±94,7 мкм.
Ширина клубочковой зоны, ответственной за секрецию минералокортикоидов,
на-против, уменьшена — 68,9 ±10,8 мкм. Через 6 ч после
однократного слабого воздействия кора надпочечников увеличена
значительно меньше (рис. 11): ширина пучковой зоны
—368,4±22,7 мкм, ширина клубочковой зоны —
84,8±9,6 мкм. Одновременно происходит увеличение ядер клеток
этих зон и изменение содержания аскорбиновой кислоты и липи-дов, РНК и
нейтральных мукополисахаридов, свидетельствующие о повышении
функцио-нальной активности этих зон коры надпочечников. По сравнению с
реакцией тревоги стресса изменяется соотношение в крови калия и натрия
в пользу натрия, что также кос-венно свидетельствует о том, что
подавления секреции минералокортикоидов не происхо-дит. Об этом же
говорит тот факт, что тимус не уменьшается.
Функция щитовидной железы в стадии ориентировки находится в
пределах нижней по-ловины зоны нормы.
Содержание в крови
22
Рис 8. Комплекс изменений в организме при
стрессе (схема). Резистентное снижена в стадиях
тревоги и истощения, повышена в стадии резистентности.
Энергетические траты высоки. Провоспалительный
по-тенциал резко повышен (в стадиях тревоги и
истощения) с угнетением активности защитных систем
белково-связанного йода (БСЙ) у крыс в стадии ориентировки составляет
6, ±20,9 мкг %; бутанол-экстрагированного йода (БЭЙ) —
4,9±0,7 мкг%; в контроле БСЙ -5,9±0,4 мкг %; БЭЙ
—3,6 ± 0,3 мкг %. У людей количество БСЙ в реакции
тренировки —
23

Рис 9 Комплекс изменении в организме
при реакции тренировки (схема) Пассивная
резистентность повышена в стадии ориентировки, активная
резистентность существенно повышена
в стадии тренированности Энергетические
траты минимальны Умеренно
повышен провоспа лительный потенциал в стадии ориентировки
без подавления за-щит ных систем организма и иммунодепрессии
7 9 ±0,35 мкг %, БЭЙ —6,4±0,28 мкг
% БСЙ в контроле —5,9± O,4 мкг
%, тироксин (Т)
63±9,1 нмоль/л, трийодтиронин —
1,5±0,36 нмоль/л.
Функция щитовидной железы при стрессе в реакции тревоги
24

Рис 10 На надпочечники крыс Реакция стресс сужение клубочковои и
расширение пучковой зоны. Окр по Пурвису и Грисбаху ув 40X7

Рис 11 Надпочечники крыс Реакция тренировки Соотношение пучковой и
кпубочковой зон соответствует норме Окр гематоксилин
эозином Ув 20X7
снижена - содержание БСЙ в крови крыс 1,7±0,3 мкг %, БЭЙ
-0,8+0,3 мкг %. У людей при развитии стресса БСЙ -3,1 ± 0,35 мкг
%, БЭЙ -1,9±0,23 мкг %, тироксин (Т) 41±10,8 нмоль/
г три
25
йодтиронин —1,1 ±0,31 нмоль/л. Морфологические
исследования показали, что в фазе ориентировки отсутствуют признаки
угнетения секреции щитовидной железы (рис. 12), в то время как при
реакции
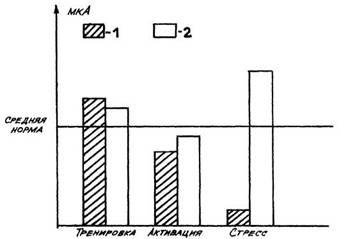
Рис. 12. Щитовидная железа крыс. Реакция тренировки: фолликулы средних
размеров, тиреоид-ный эпите-лий кубический. Окр. гематоксилин-эозином.
Ув. 40X7
тревоги тиреоидный эпителий уплощен, имеются явления карио-лизиса,
сгущенный кол-лоид окрашивается интенсивно, во многих фолликулах
эпителий десквамирован, отмеча-ется слияние фолликулов и увеличение их
размеров, т. е. и морфологическая картина го-ворит о снижении
функциональной активности щитовидной железы в первую фазу стрес-са
(рис. 13).
В стадии ориентировки у крыс продолжительность активных фаз полового
цикла — в пределах нормы, гонадотропная активность гипофиза
— в пределах нижней полови-ны зоны нормы, что говорит об
умеренной, нормальной активности половых желез. Функция половых желез в
реакции тревоги снижается: снижена гонадотропная активность гипофиза,
нарушается половой цикл либо с преобладанием диэструса, либо с резким
пре-обладанием активных фаз цикла. Морфологические исследования
семенников в стадии ориентировки показали отсутствие признаков
угнетения сперматогенеза (рис. 14), в то время как при реакции тревоги
в большинстве канальцев имело место угнетение сперма-тогенеза (рис. 15).
В гипофизе наблюдалась тенденция к увеличению числа тирео-
26
Рис. 13. Щитовидная железа крыс. Реакция стресс: увеличение размеров фолликулов.
уплощение тиреоидного эпителия. Окр. гематоксилин-эозином. Ув. 40Х 7
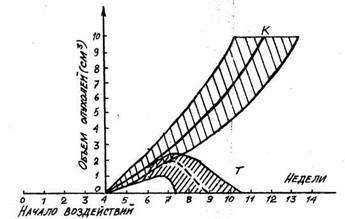
Рис. 14. Семенники крыс. Реакция тренировки. Количество сперматозоидов
в семенных канальцах в пределах нормы. Окр. по Эйнарсону. Ув. 40Х 10
трофов с признаками увеличения в них секреции — гипертрофия
клеток, вакуолизация цитоплаз-мы, увеличение числа гонадотрофов, в то
время как при реакции тревоги стресса — уменьшение чи^ля
гонадотрофов и увеличение числа ацидофилов (Гаркави Л. X., 1968. 1969;
Уколова М. А., Бордюшков Ю. Н., Гашникова Л. И., Квакина Е. В., 1972).
При развитии реакции тревоги стресса отмечается повышение
коагуляционного потенциа-ла: укорочено время реакции «R»
(5,5 ± ±0,6 по сравнению с 11,0±1,7 в контроле), в
результате чего укорачивается общее время свертывания «Т»
(98,0±8,6 по сравнению с 124,7±4,0 в контроле). В фазе
ориентировки реакции тренировки со стороны свертывающей системы крови
отмечается противоположная стрессу ответная реакция —
гипокоагуляция: более чем вдвое удлиняется время образования сгустка
«К» (44,1 ±:7,5 по сравнению с 20Д±2,6 в
контроле). Так как время реакции «R» при этом существенно
не изменяется, можно предполагать, что удлинение «R + +
К» (52,8±8,6 по сравнению с 31,1 ±2,03 в
контроле) происходит
27
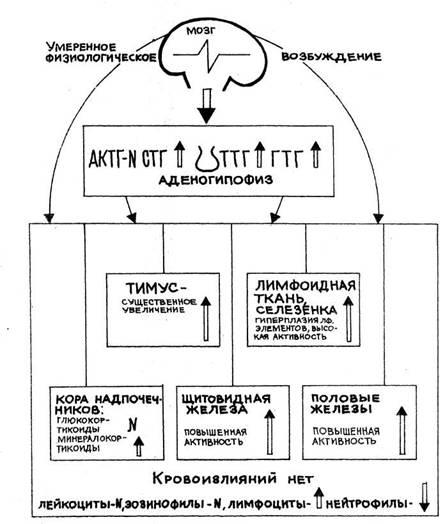
Рис 15 Семенники крыс Реакция стресс уменьшение количе ства
сперматозоидов в семен-нык канальцах Окр по Эйнарсону .
Ув. 40X 10
за счет усиления активности противосвертывающей системы. Происходит
также сниже-ние констант фибриногена Ма и Е (Уколова М. А., Гаркави Л
X., Квакина Е. В., 1969; Гар-кави Л. X , Уколова М. А , Григорьева Е. Г
, 1971; Григорьева Е. Г , 1975; Григорьева Е. Г.. Гаркави Л. X ,
Уколова М. А., 1980).
Изменения метаболизма в стадии ориентировки отличны от изменений при
стрессе. При исследовании стадии тревоги стресса было получено резкое
повышение (в 7—8 раз) содержания аммиака, повышение (на
60—90%) глютамина и снижение (вдвое) глютами-новой кислоты.
Интенсивность тканевого дыхании в мозгу не изменялась Резко повыша-лось
накопление макроэргических фосфатов (АТФ + КФ), эстерификация
неорганическо-го фосфора повышалась на 30%. Анаэробный и особенно
аэробный гликолиз резко возра-стали соответственно на 60 — 70% и
в 2-2,5 раза), создавалось состояние напряжения ме-таболизма.
Изучение энергетического обмена на уровне митохондрий покатало, что при
стрес-се были значительно снижены фонд эндогенной янтарной кислоты
(ЭЯК) и ее воспроиз-водство, выявлялось глубокое щавелево-уксусное (ЩО)
ограничение активности СДГ, снижающее избыточную скорость субстратного
окисления. При более тяжком стрессе скорости окисления существенно
снижены, время фосфорилирования увеличено, фонд ЭЯК довольно высок.
В этом
28
случае происходит переключение с углеводного на липидный источник
энергии (Барсуко-ва Л. П., Гаркави Л. X., Котляревская Е. С.
Марьяновская Г. Я., 1983; Гаркави Л. X., Квакина
Е. Б. Марья новская Г. Я. Барсукова Л. П.,
Котляревская Е. С, 1984). Содержа-ние РНК существенно
снижалось: в коре головного мозга на 55
-60%, в надпочечниках — на 20 — 25%. Еще более резко cнижа
лось содержание белка: в мозгу — на 55—65% (в
основном за счет резкою снижения альбуминов), в печени
— на 40- 45%, в селезенке — на 40 —45%, в семенниках
— на 50—55%. В сыворотке крови отмечалось
небольшое снижение общего белка
— на 10—15%., в основном за
счет альбуминов (Гаркави Л. Х.
1969). Происходил резкий подъем
активности ацетилхолинэстеразы в коре
го-ловного мозга и гипоталамуса (в 3 — 4 раза) и крови (в 2
— 3 раза) (Арефьева Л. М , 1972) При
реакции тренировки в мозгу наблюдается небольшое нарастание содержания
аммиака (примерно в 1,5 раза) и небольшое снижение глютамина (на 25
— 30%) при от-сутствии изменений глютаминовой кислоты (Гаркави Л.
Х. Рогозная А. В , 1963; Гаркави Л. X . 1969). Интенсивность тканевого
дыхания и эстерификации неорганического фосфо-ра существенно не
изменяется, отмечается некоторое
уменьшение накопления
мак-роэргических фосфатов (АТФ+КФ) — на
30—40%, снижение анаэробного гликолиза - на 20—25% и
аэробного гликолиза - также на 20—25% (Гаркави Л.Х. , 1969). При
реакции тренировки снижаются и процессы катаболизма и процессы
анаболизма: высокий фонд ЭЯК, скорости окисления субстратов и
окислительного фосфорилирования, время фосфо-рилирования —
умеренны. Количество РНК снижается в коре головного мозга на 25
—30%, в гипоталамусе — на 30—35%, в печени
- на 25-30%, в тимусе — на 10—15%
(Гаркави Л. X,, Трапезонцева Р А. , 1964). Происходит
снижение содержания общего белка в мозгу на 30—40% за счет
снижения глобулинов (?1—40 — 50%, ? 2
— 40 -50%, ?—30-40% и ? —
40—50%). В печени снижение общего
белка на 15—20% происходит за счет всех белковых фракций, в
селезенке — на 20—30%, главным образом, за счет
сни-жения ? -глобулинов, в семеннике — на 20— 30% за
счет снижения ?1 и ?2-глобулинов. В сыворотке крови также отмечалось
снижение общего белка на 7 -10% за счет умеренного снижения всех
фракций (Гаркави Л. Х , 1969). Отмечалось
небольшое повышение ак-тивности ацетилхолинэстеразы в коре
головного мозга (на 20 — 30%), уменьшение - в
гипоталамусе (на 20—25%) при отсутствии существенных
изменений в крови (Квакина Е. Б., Уколова М. А., Гаркави Л. X.,
Арефьева Л. М., 1971).
Изменения, характерные для стадии ориентировки реакции тренировки,
законо-мерно возникают уже через 6 ч после действия любого по качеству,
но слабого раздражи-теля (электрическое раздражение гипоталамуса,
магнитное поле, нейротропные вещества и т.п.
29
Что же происходит в ЦНС — системе, формирующей развитие любой
адаптацион-ной реакции? Исследования показали, что в ЦНС в стадии
ориентировки преобладает со-стояние охранительного торможения.
Возбудимость нервных структур гипоталамуса сни-жена на
32±4% (рис. 16) по сравнению с исходной
(Квакина Е. Б., Ква-
Рис 16. Изменение возбудимости в первые стадии реакции тренировки
и стресса (по величине порогов электрического раздражения) через 1 час
после воздействия (1) и через 24 часа после воздействия (2)
кин С. Д., Котляревская Е. С, 1969; Квакина Е. Б. и др., 1969; Квакина
Е. В., 1972; Котля-ревская Е. С, 1974). В ЭЭГ отмечалось увеличение
числа медленных волн (дельта-волн). Это соответствует представлениям П.
С. Симонова (1962) о развитии в ЦНС при действии слабого стимула
превентивного торможения. В реакции тревоги стресса удалось отметить в
ЭЭГ отсутствие регулярности смены ритмов и низкую амплитуду колебаний.
Резкое возрастание возбудимости (снижение порогов на 70—90%)
сменялось ее резким снижени-ем (повышение порогов на 70—90%, см.
рис. 16). Изучение сверхмедленных колебаний потенциалов головного мозга
(СМКП), описанных впервые Н. А. Аладжаловой (1962), по-казало, что в
стадии ориентировки отмечается высокая амплитуда волн (до 40—60
мВ) при преобладании волн с большим периодом (0,5 — 1,5 ч), в то
время ?как в реакции тре-воги отмечается низкая амплитуда волн
(5—25 мВ) при преобладании волн с малым пе-риодом (17—30
мин). Необходимо отметить, что постоянное магнитное поле (ПМП) даже
разной индукции вызывает преимущественно развитие реакции тренировки,
что мы свя-зываем с его тормозным действием (Холодов Ю. А., 1966—
1985). С помощью ПМП ре-акция тренировки вызывалась не только
30
нами, но и другими авторами (Шишло М. А., Кубли С. X., Шимкевич Л. Л., 1981).
Поскольку развившееся в стадии ориентировки охранительное торможение
умень-шает чувствительность, то если действующий фактор остается в
дальнейшем таким же слабым, организм перестает на него реагировать. С
помощью торможения организм за-щищает себя от бесчисленного множества
разнообразных, повторяющихся без изменений интенсивности, слабых
раздражителей, на которые можно не реагировать. Развившееся в конце
стадии тревоги стресса запредельное торможение резко снижает
чувствительность, и если величина действующего стрессорного агента не
увеличится, то повторное действие такого же раздражителя уже не
вызывает стресса. Таким образом, организм защищен тор-можением с двух
сторон: от действия малых, несущественных раздражителей —
первич-ным охранительным торможением; от перераздражения, истощения и
гибели — вторич-ным запредельным торможением (Симонов П. В.,
1962; Гаркави Л. X., Квакина Е. Б., Уко-лова М. А., 1979).
Для поддерживания в организме реакции тренировки нужно специально
немного, причем волнообразно, повышать интенсивность раздражителя,
чтобы он снова достиг ми-нимальной величины, вызывающей общую реакцию
организма. При этом будет разви-ваться следующая, вторая стадия реакции
тренировки — стадия «перестройки». Сущность
перестройки заключается в постепенном снижении секреции
глюкокортикоидных гормо-нов и постепенном повышении секреции
минералокортикоидных гормонов. Активность тимико-лимфатической системы
и системы соединительной ткачи также постепенно по-вышается, т. е.
активность защитных систем организма при каждодневных целенаправ-ленных
тренировочных воздействиях медленно и постепенно нарастает.
Этот процесс повышения активности защитных систем путем тренировки идет
волнообразно, а не нарастает прямолинейно. Так, тимус то несколько
уменьшается (до Нижних границ контроля), то вновь увеличивается, В
головном мозгу после каждого воз-действия какое-то время преобладает
состояние охранительного торможения, т. е. идет как бы тренировка
торможением, и абсолютная величина минимального раздражителя,
вызывающего эту реакцию, постепенно возрастает.
В метаболизме также происходят волнообразные колебания, ведущие к
снижению уровня обмена до очень низкого, энергетически выгодного уровня
с минимальным коли-чеством расходования пластических веществ.
Постепенно накапливается пластический материал — аминокислоты,
белки. После стадии «перестройки», которая длится месяц, а
иногда и более, развивается третья стадия реакции тренировки —
стадия «тренированно-сти». Тимус увеличен несколько больше,
чем в первую стадию — в 1,5-1,8 раза. Пара-метры белой
31
крови существенно не изменяются по сравнению с первой
стадией, однако в
подав-ляющем большинстве случаев
число лимфоцитов находится вблизи верхней
грани-цы, характерной для этой реакции, а число
сегментоядерных нейтрофилов — вблизи нижней границы.
Содержание глюкокортикоидов снижается до уровня средней
нормы, содержание
минералокортикоидов несколько повышено —
приближается к верхней по-ловине зоны нормы. В коре надпочечников
отмечается расширение клубочковой
зоны с признаками
высокой активности.
Функциональная активность
щитовидной железы остается умеренно
повышенной: увеличены высота и размер ядер
тиреоидного эпителия, отмечается вакуолизация коллоида,
Содержание в крови БСЙ, БЭЙ, тироксина и трийодтиронина близко
к исходному уровню или несколько повышено.
Активность половых желез остается в пределах нормы. В гипофизе число
гонадотрофов и тиреотро-фов увеличено, а число ацидофилов не изменено.
Система свертывания крови характеризуется гипокоагуляционным сдвигом,
не-сколько менее выраженным, чем в стадии ориентировки. В мозгу
снижается содержание аммиака, которое в предыдущих стадиях этой реакции
было несколько повышено (на 15—20%), содержание глютамина также
умеренно снижено — на 25 — 30%. Содержание глю-таминовой
кислоты — без изменений. Уровень белка в мозгу и других органах
несколько увеличен (на 10—20%). Повышено количество РНК в мозгу
(на 35—40%). Повышено со-держание белка в коре головного мозга
(на 10—15%) и в гипоталамусе (на I 15 — 20%). Это говорит о
преобладании процессов анаболизма с накоплением пластического
мате-риала. В ЦНС в этой стадии также преобладает процесс
охранительного торможения (Гаркави Л. X., 1969; Гаркави Л. X.,. Квакина
Е. В., Уколова М. А., 1975).
Резистентность организма при развитии реакции
тренировки
Систематическое повторение слабых, но постепенно нарастающих
раздражителей с течением времени приводит к повышению активности
защитных систем, в связи с чем и организм становится устойчивым к
повреждающим воздействиям уже не только за счет развития торможения в
головном мозгу, но и благодаря повышению активности защитных систем.
Особенно интересно то, что каким бы воздействием не
«тренировали», повыше-ние устойчивости происходит не только
к этому воздействию, но и к другим. Факт повы-шения неспецифической
резистентности организма при тренировке различными раздражителями
неоднократно отмечается в литературе.
Селье и его последователи установили, что неспецифическая
резистентность орга-низма повышена только в кратковременной стадии
резистентности стресса и
достига-ется «дорогой ценой»
— ценой
32
повреждения и больших энергетических трат. Это плата за возможность
защитить себя от непосредственной угрозы, от действия сильного,
повреждающего, а часто и патогенного фактора. В стадии реакции тревоги
и особенно в стадии истощения резистентность организма снижена (рис.
17).
Рис. 17. Схема изменения резистентности
организма: а — при стрессе, б— при
реакции
тренировки
Как изменяется уровень резистентности при реакции тренировки, в ее различных стадиях?
В первой стадии реакции тренировки — ориентировки — про
исходит уже некото-рое повышение неспецифической резистентности
организма. Это показано нами в опытах по повышению резистентности к
повреждающему действию противоопухолевых химиоп-репаратов и облучения в
так называемых терапевтических дозах. Кок известно, и химиоп-репараты,
и облучение оказывают повреждающее действие на организм, вызывая
измене-ния по типу стресса. Особенно поражается аппарат кроветворения.
Электрическое раз-драженно гипоталамуса током слабой силы, постоянное
магнитное поле, адреналин в ма-лых дозах (перед облучением) оказывали
защитное действие при разовом облучении бе-лых крыс в дозе 6 Гр,
уменьшая лейкопению и летальность. При локальном однократном облучении
области таза крыс в дозе 20 Гр (100%-я летальность) после тренировочных
воздействий летальность снижалась до 40—60%. При облучении
туловища крыс в дозе 7 Гр число лейкоцитов было снижено в течение трех
недель после облучения (4250 ± 300, 5920 ± 420,
6920 ± 300 лейкоцитов соответственно через 1, 2, 3 недели), в то
время как в под опытной группе лейкопения была только в первую неделю
(5400 ± 560), после чего наступало быстрое восстановление числа
лейкоцитов (10250 ± 1000 и 11120 ± 500 через 2 и 3
недели) (Комурджиев X. А. и др., 1967; Квакина Е. Б., 1972).
При введении циклофосфана в дозе, в 3 раза превышающей 100%-ю летальную
(400 мг/кг), тренировочное воздействие хотя и не снижало летальность,
но уменьшало лейкопению (через 2 дня число лейкоцитов в опытной группе
упало на 115004 ± 3100, в контрольной —
33
на 20900±2300) (Уколова М. А., Квакина Е. В., Рудой Ф. И.,
1969). Введение циклофана, как дробное, в дозе 7 —14 мг/кг,
суммарно — 190 мг/кг, так и однократное, в дозе 90 мг/кг,
приводило к развитию у крыс стресса с лейкопенией и к гибели в половине
случаев. Специальное получение у животных подопытных групп реакции
тренировки при помощи ежедневных воздействий постоянного МП 30 мТл и
250 мТл при тех же условиях введе-ния циклофосфана приводило к
существенному снижению летальности и уменьшению лейкопении. Так, через
2 дня после инъекции циклофосфана в дозе 90 мг/кг число лейко-цитов в
контрольной группе было 1060±521, в
подопытной—3049±768 (Р<0,001) при оди-наковом исходном
количестве (Квакина Е. Б., Гаркави Л. X., Уколова М. А.,
Рудой-Тарасьянц Ф. М., Мулатова А. К., Григорьева Е. Г., 1980).
Исследовалось также влияние первой стадии реакции тренировки на рост
опухо-лей. Противоопухолевого эффекта получить не удалось. Для
сравнения при этих же по-вреждающих агентах вызывали развитие стресса.
Это приводило лишь к усилению повре-ждающего действия лучевой и
химиотерапии и ускорению роста опухолей.
С чем связано повышение резистентности при развитии первой стадии
реакции тренировки? Гормональные изменения, как уже говорилось, таковы,
что они не приводят к существенному повышению активности защитных
систем организма. По нашему мне-нию, в этой стадии повышение
резистентности происходит за счет того, что в ЦНС преоб-ладает
состояние охранительного торможения, coпровождающееся снижением
возбуди-мости, благодаря чему организм становится менее чувствительным,
т. е. резистентность в стадии ориентировки можно назвать пассивной. Мы
думаем, что повышение резистентно-сти при стрессе также во многом
связано с развитием торможения, но не охранительного, а запредельного,
тоже снижающего возбудимость ЦНС и чувствительность организма.
Необходимо отметить, что в стадии ориентировки происходит, хотя и
меньшее чем при реакции тревоги, повышение секреции глюкокортикоидов.
Поэтому в этой стадии ре-акция тренировки обладает мягким
противовоспалительным действием, не таким силь-ным, как стресс, но без
каких-либо элементов повреждения защитных систем организма. Повреждение
защитных систем, иммунодепрессия, как известно, является нежелательным
последствием применения глюкокортикоидов с противовоспалительной целью
в дозах, вызывающих стресс.
В фазе тренированности наряду с повышением пассивной резистентности
происхо-дит подъем резистентности за счет истинного возрастания
активности защитных систем организма, т. е. повышается и активная
резистентность. В этой стадии наблюдалось за-щитное влияние по
отношению к многократному (дробному) облучению и многократному введению
токсических препаратов.
34
При локальном облучении крыс (область таза) в суммарной дозе 20 Гр по 4
Гр за сеанс в подопытных группах лейкопения
отсутствовала.
При введении циклофосфана в суммарной дозе 147 мг/кг (по 7 — 14
мг/кг на инъ-екцию) в контрольной группе происходила гибель 50%
животных, в то время как у под-опытных с развитием стадии
тренированности выживало 90—95% животных. Аналогич-ные результаты
были получены по отношению к тиоТЭФу и сарколизину (Квакина Е. Б.,
1972; Квакина Е. В., Гаркави Л. X., Уколова М. А. Мулатова А. К.,
Григорьева Е. Г., 1980; Захарюта Ф. М., 1989),
В стадии тренированности удалось получить повышение противоопухолевой
рези-стентности (Гаркави Л. X., 1962; Уколова М. А... Бордюшков Ю. Н.,
Гаркави Л. X., Ква-кина Е. Б. и др., 1963: Uko lova М. A., Bordjushkov
Y. N., Garkavi L. Н., Goncharova V. К., Kva-kina E. В., 1964). С
помощью многократно повторяющихся слабых воздействий дос-тигалась
стадия тренированности, после чего животным перевивали различные
опухоли (экспериментальные саркомы и рак молочной железы). Вначале
опухоли: росли, как и в контроле, а затем рост их начинал тормозиться и
происходило либо отставание в росте от опухолей контрольных животных,
либо полная регрессия в то время, как в контроле они продолжали расти и
приводили животных к гибели (рис.
18). Подобное влияние ока-зывали слабые

Рис. 18. Рост экспериментальных опухолей: К — без воздействия, Т — при тренировочных воздействиях на организм
повторные воздействия и на опухоли, развивающиеся в организме из его
нормальных тка-ней,— опухоли яичника, получаемые по методике
Biskind G. и Biskind M. (1949) из транс-плантатов яичника в селезенке
кастрированных крыс. Происходило торможение роста
35
и полное рассасывание трансплантатов, что предотвращало развитие из них
злокачествен-ных опухолей (Уколова М. А., Бордюшков Ю. Н., Гаркави Л.
X., Квакина Е. Б. и др., 1963; Уколова М. А., 1972).
Такой профилактический характер противоопухолевого влияния реакции
трени-ровки сходен с характером действия на опухолевый процесс
адаптогенов. Наиболее чет-кое противоопухолевое влияние адаптогены
оказывали именно при предварительном применении, и Н. В. Лазарев (1968)
особенно подчеркивал их роль в профилактике опухо-левой болезни.
По-видимому, чаще всего адаптогены применялись в дозах, вызывающих
развитие реакции тренировки. Воздействия на уже выросшие
экспериментальные опухо-ли, приводящие в развитию реакции тренировки,
вызывали лишь торможение их роста (Гаркави Л. X., Квакина Е. Б.,
Мулатова А. К., Шихля-рова А. И., 1980).
Характеристика стадий реакции тренировки, влияние на
противовоспалительный потенциал организма, на уровень его
резистентности и небольшая «цена» резистентности —
все это говорит о том, что реакцию тренировки целесообразно
использовать на практи-ке. Эта реакция может быть применена для мягкого
противовоспалительного действия. Нельзя, однако, забывать, что
поддержание высокой резистентности при реакции трени-ровки возможно
только при систематическом повторении тренировочных воздействий
(упражнений). Прекращение таких воздействий приводит к быстрому
снижению рези-стентности — детренированности. По нашему мнению,
это связано с низкими энергетиче-скими тратами при реакции тренировки.
Переход в любую другую реакцию предъявляет к организму как бы большие
требования.
Благодаря способности быстро повышать пассивную резистентность,
оказывать мягкое противовоспалительное влияние, защищать организм от
повреждающего действия лекарственной и лучевой терапии (особенно при
выраженной лейкопении) и свойственной этой реакции экономичности
энергозатрат, реакция тренировки может найти лечебное применение,
особенно в комплексе с другими терапевтическими мероприятиями (Гаркави
Л. X., Квакина Е. Б., Уколова М. А., 1979).
РЕАКЦИЯ АКТИВАЦИИ — ОБЩАЯ НЕСПЕЦИФИЧЕСКАЯ АДАПТАЦИОННАЯ РЕАКЦИЯ
НА РАЗДРАЖИТЕЛИ «СРЕДНЕЙ» СИЛЫ
Как уже отмечалось, адаптация к сильным раздражителям (реакция стресс)
обеспе-чивается не только путем чрезмерной стимуляции одних систем
организма, но и путем подавления активности других — защитных
систем. При адаптации к слабым раздражите-
36
лям (реакция тренировки) не происходит ни подавления, ни существенной
стимуляции защитных систем. Существует ли реакция, при которой
активность защитных систем ор-ганизма целесообразно повышать, т. е.
реакция, несущая основную антистрессорную функцию? Исследования
показали, что это реакция на раздражители, промежуточные ме-жду
сильными и слабыми, так называемой средней силы. Эти раздражители не
такие сильные, чтобы было целесообразно подавлять защитные системы для
снижения реакции на них. Вместе с тем они и не слабые. Чтобы
приспособиться к действию такого раздра-жителя, необходимо быстро
повысить активность защитных
систем.
В результате исследований было установлено, что в ответ на действие
различных по качеству раздражителей средней силы развивается общая
неспецифическая адаптаци-онная реакция, названная «реакцией
активации» (Гаркави Л. X., 1968, 1969), так как она
характеризуется быстрым подъемом активности защитных и регуляторных
систем орга-низма.
Реакции активации так же, как реакции стресс и реакции тренировки,
свойствен свой комплекс изменений в нейроэндокринной системе (рис. 19).
Эта реакция также про-текает стадийно, и каждая ее стадия
характеризуется определенным уровнем неспецифи-ческой резистентности
организма.
Чем характеризуется реакция активации? Как она развивается?
Средние раздражители также довольно часто встречаются в природе и
повседнев-ной жизни. Первая стадия в реакции на раздражители средней
силы, названная стадией первичной активации, так же, как и первые
стадии реакции тренировки и стресса, форми-руется через 6 ч и держится
24—48 ч после воздействия. Однако характер изменений от-личается
как от изменений в стадии ориентировки реакции тренировки, так и от
измене-ний в реакции тревоги стресса. Так, в стадии первичной активации
тимус существенно увеличен — в 2-2,5 раза по сравнению с
контролем (статистически значимо больше, чем в стадии ориентировки).
Его величина находится.в пределах верхней половины зоны нор-мы.
Гистологическое исследование показало, что это истинная гипертрофия
тимуса (рис. 20). Отмечались гиперплазия лимфоидной ткани, неравномерно
выраженная в различных дольках, и некоторая гипертрофия долек. Зона
коркового вещества расширена, местами в виде тяжей вдается в мозговое
вещество. Была отмечена гипертрофия лимфатических уз-лов. Структура их
значительно стерто вследствие гиперплазии лимфоидных элементов.
Наблюдались гиперплазия и увеличение размеров клеток ретикулоэндотелия.
В некоторых случаях была выражена плазматизация лимфоидных эле ментов.
В лимфатиче-ских узлах крыс, находившихся в реакции активации,
вследствие повышения митотиче-ской активности пиронинофильных
широкоплазменных лимфоцитов, утолщаются мякот-
37
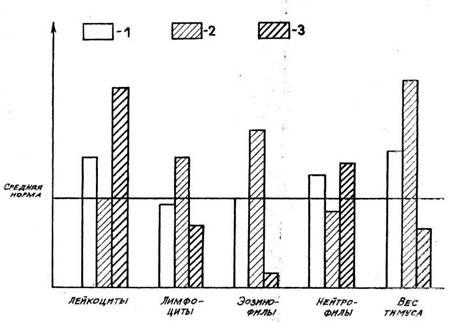
Рис. 19. Комплекс изменений в организме при реакции активации (схема).
Активная резистентность сущест-венно повышена в стадии первичной
активации и особенно — в стадии стойкой активации. Энергетические
траты умеренны. Существенно повышен провоспалительный потенциал
организма
ные тяжи и усиливается дифференцировка пиронинофильных лимфоцитов в
зрелые формы (рис. 21). В селезенке происходит увеличение размеров ядер
ретикулярных клеток и формирование из них мегакариоцитов,
количество которых повышено.
Герминативные
38
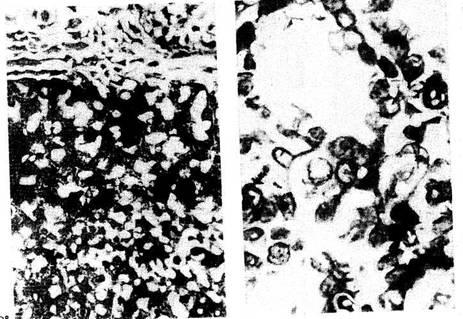
Рис. 20. Тимус крыс. Реакция активации: внедрение тяжей лимфоидной
ткани из корковой части в мозговую при расширении корковой части. Окр.
но Браше в модификации А. А. Симаковой. Ув. 8X7
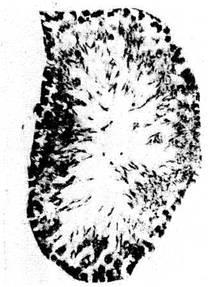
Рис. 21. Лимфоузлы крыс, реакция активации: а — утолщение
мякотных тяжей и дифференцировка клеточных элементов в зрелые лимфоциты
в единичных мякотных тяжах. Окр. по Браше в модификации А. А.
Симаковой. Ув. 16X7;
б — плазматизация клеточных элементов в мякотных тяжах. Окр. по Браше в модификации А. А. Симаковой. Ув. 40X7
39
центры в лимфоидных узелках состоят преимущественно из лимфобластов,
больших и средних лимфоцитов. По сравнению с реакцией тренировки
изменяется соотношение раз-меров герминативных центров и пояса короны
малых лимфоцитов в пользу последних (рис. 22). Все
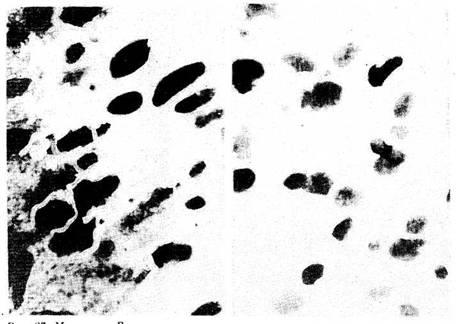
Рис. 22. Селезенка крыс. Реакция активации: некоторое сужение
герминативных центров и расширение среднего пояса фолликулов. Окр. по
Браше в модификации А. А. Симаковой. Ув. 16X7
это говорит о физиологической стимуляции тимико-лимфатической системы
(Гаркави Л. X., 1968, 1969; Квакина Е. Б., 1972; Мулатова А. К., 1983).
Стадию первичной активации так же, как и стадию ориентировки и реакцию
тревоги, ха-рактеризуют определенные соотношения форменных элементов
белой крови (рис. 23). В стадии первичной активации у крыс число
лимфоцитов — в пределах верхней половины зоны нормы и выше
(60—80%), сегментоядерных нейтрофилов — в пределах нижней
половины зоны нормы и ниже, число лейкоцитов, эозинофилов,
палочкоядерных нейтрофилов и моноцитов — в пределах нормы.
У человека параметры реакции активации следующие: число лимфоцитов
— в пределах верхней половины зоны нормы и несколько выше (от 28
до 40—45%—индивидуально), сегмен-тоядерных нейтрофилов
— в пределах нижней половины зоны нормы и ниже, число лейкоцитов
— в пределах нормы (4,0—8,0) -109, число
эозинофилов, палочкоядерных
нейтрофилов, моно-цитов также
в норме
40
Рис: 23. Лейкоциты, лейкоцитарная формула и масса тимуса в первые стадии реакции тренировки (1), акти-вации (2) и стресса (3)
(Гаркави Л. X., 1968, 1969; Гаркави Л. X, Гарибьян В. Р., 1968; Квакина
Е. В., 1972; Гарка-ви Л. X., Квакина Е. В., Уколова М. Л., 1974, 1975).
При реакции активации в лимфоцитах значительно повышена активность СДГ
и несколько снижена — ЛДГ. В стадии первичной активации
увеличивается секреция минералокортикоидных гормонов коры
надпочечни-ков, секреция же глюкокортикоидных гормонов находится в
пределах нормы. Секреция минералокортикоидов преобладает. Через 6 ч
после однократного воздействия средней силы (дозы) у людей содержание
17-ОКС в крови —10,0 ± 0,6 мкг%, 11-ОКС у крыс —35,7
± 2,4 мкг%, кортизола в крови людей—249 ± 68,5
нмоль/л.
Морфофункциональное исследование коры надпочечников (Гаркави Л. X.,
Квакина Е. В., Уколова М. А., 1979; Шихлярова А. И., 1985) показало,
что кора надпочечников увеличена, в основном, за счет клубочковой зоны
(130,154±24,0 мкм по сравнению с 75,954 ±23,34 мкм),
ответственной за секрецию минералокортикоидов (рис. 24), а пучко-вая
зона несколько уменьшена (195,70+32,55 мкм по сравнению с
397,70± 120,27 мкм). Отмечалось увеличение ядер клеток
клубочковой зоны. Наблюдались признаки повыше-ния функциональной
активности в виде вакуолизации цитоплазмы клеток клубочковой и пучковой
зоны и вакуолизация цитоплазмы клеток
41
мозговой части. Отношение калия к натрию в крови по сравнению с
реакцией тревоги из-меняется в сторону преобладания натрия, т. е.
остается таким же, как в первой стадии ре-акции тренировки, но при
более высоком уровне в крови и калия, и натрия (Гаркави Л. X. и др.,
1968; Гаркави Л. X., 1969; Бордюшков Ю. Н., Гашникова Л. И., Квакииа Е.
В., 1971; Квакина Е. В., 1972). Все это, а также увеличение тимуса и
лимфоцитоз свидетельствуют о преобладании минералокортикоидной
функции коры надпочечников
над глюко-
кортикоидной.
Функция щитовидной железы повышается, однако патологических проявлений
гиперти-реоза нет. Содержание БСЙ у крыс — 9,84±0,7 мкг %,
БЭЙ—8,74±0,4 мкг %. У людей БСЙ составляет 9,3 ±
0,5 мкг %, БЭЙ —7,8±0,35% мкг %, тироксин
—63± ±9,1 нмоль/л, трийодтиронин
—1,5±0,36 нмоль/л. Морфологические исследования показали,
что в щи-товидной железе эпителий имеет цилиндрическую или кубическую
форму, отмечается увели-

Рис. 24. Надпочечники крыс. Реакция активации: расширение клубочковой
зоны, увеличение содержания липидов в цитоплазме клеток. Окр. Суданом
III. Ув. 20X7
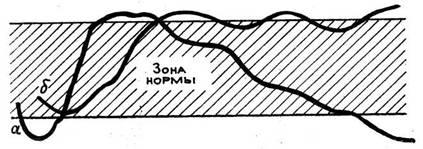
Рис. 25. Щитовидная железа крыс. Реакция активации: фолликулы средних
размеров: тиреоидный эпителий высокий, цилиндрический, резко выражена
вакуолизация коллоида. Окр. гематоксилин-эозином. Ув. 40Х 7
42
чение ядер тиреоцитов, количества делящихся клеток, их размеров (рис.
25). В большин-стве фолликулов коллоид жидкий, окрашивается слабо.
Исследование величины синтети-ческой активности клеток щитовидной
железы методом микроспектральной люминесценции (Карнаухов В. Н., 1978)
выявило ее существенное увеличение при реакции акти-вации и снижение
при стрессе. Так, величина а (представляющая отношение одно- и
дву-спиральных нуклеиновых кислот) при реакции активации составляет
0,1-0,5, а при стрессе 0,03-0,2, Р<0,01 (Гаркави Л. X., Квакина Е.
В., Шей-ко Е. А., 1987).
Умеренно повышается активность половых желез: несколько увеличивается
продолжи-тельность активных фаз полового цикла, повышается
гонадотропная активность гипофиза. В семенниках на блюдается разная
степень активации сперматогенеза: в пресперма-тидах — большое
количество фигур митоза; в клетках Сертоли —
сперматозоидов (рис. 26). В гипофизе —
увеличение числа тирео-
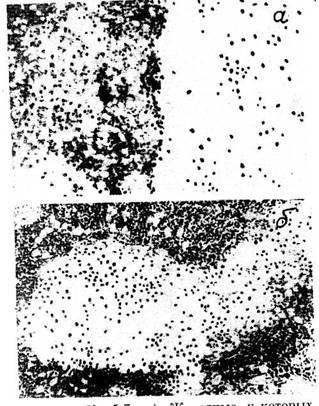
Рис. 26. Семенники крыс. Реакция активации: значительное увеличение
количества сперматозоидов в се-менных канальцах (поперечный срез). Окр.
по Эйнарсону. У в. 40X10
трофов и гонадотрофов с признаками увеличения секреции —
гипертрофия клеток, вакуолизация цитоплазмы (Гаркави Л. X., 1969;
Бордюшков Ю. Н., Гашникова Л. И., Квакина Е. В., 1971; Квакина Е. П.,
1972; Гаркави Л. X., Квакина Е. В., Уколова М. А., 1975).
При развитии стадии первичной активации происходит уменьшение времени
реакции «R» (7,0±0,35 мм по сравнению с
11,04± ±1,7 мм в контроле), не отражающееся на общем
вре-мени свертывания Т, которое остается неизменным, т. е. функции
свертывающей
43
и антисвертывающей системы уравновешены (Гаркави Л. X., Уколова М. А.,
Григорьева Е. Г., 1971; Квакина Е. Б., Григорьева Е. Г., Марьяновская
Г. Я., 1971; Григорьева Е. Г., Гаркави Л. X., Уколова М. А.,
1980).
Закономерно изменяются некоторые
показатели метаболизма, В мозгу проис-ходит
увеличение содержания аммиака в 2,5 раза при
отсутствии изменений в содер-жании
глютамина и глютаминовой кислоты (Архангельская А. В.
и др., 1968). Тканевое дыхание не- сколько повышено (на
15—20%), а эстерификация неорганического фос-фора не
изменена, т. е. можно предполагать, что имеется некоторое разобщение
процессов окисления и фосфорилирования. Анаэробный и аэробный гликолиз
повышены соответст-венно на 15—20% и
75—85% (Гаркави Л. X. и др., 1969). При реакции
активации энер-гетический баланс несколько сдвигается в
сторону ускорения окислительных процессов. Возрастают
скорости окисления субстратов,
скорости окислительного
фосфорилиро-вания, сокращается время
фосфорилирования и снижается фонд эндогенной янтарной
кислоты (ЭЯК). Снижение фонда основного субстрата
окисления компенсируется воз-растанием
воспроизводства янтарной кислоты,
иногда выявляется
неглубокое ща-велево-уксусное
ограничение активности СДГ,
уменьшающее скорости окисления
субстратов (Барсукова Л. П., Гаркави Л. X., Котляревская Е. С,
Марьяновская Г. Я., 1983; Гаркави Л. X., Квакина Е. Б., Марьяновская Г.
Я., Барсукова Л. П., Котляревская Е. С, 1984).
При реакции активации происходит переход дыхательной цепи
митохондрий на преимущественное окисление
янтарной кислоты. Наблюдается интенсификация глюко-неогенеза,
несмотря на отсутствие избытка глюкокортикоидов. М. Н. Кондрашова
(1980) считает реакцию активации по состоянию энергетики
«золотой серединой», усиливающей работу по поддержанию
«неравновесного состояния» (Бауэр Э. С, 1935).
В коре головного мозга возрастает активность ацетилхолинэстеразы
— на 40 — 60% (Арефьева Л. М., 1972).
Содержание РНК повышается в большинстве тканей: в ги-поталамусе
(на 12—15%), в печени (на 11 —14%),
в тимусе (на 60—65%), в надпо-чечниках
(на 50—55%); в коре головного мозга оно не изменяется
(Гаркави Л. X., Стар-кова А. А., Якунина Н. Н., 1968). Имеется
тенденция к повышению содержания общего белка преимущественно за счет
альбуминов в тканях мозга, печени, селезенки, семенни-ков и в сыворотке
крови (Гаркави Л. X., 1969). В сыворотке и эритроцитах
увеличивается количество свободных
сульфгидрильных групп (Квакина Е. Б., Рудой Ф.
М., 1974).
Все эти изменения закономерно возникают уже через 6 ч после действия
любого по каче-ству раздражителя средней силы (электрического
раздражения гипоталамуса, действия переменного магнит-
44
ного поля, введения нейротропных
веществ, приема биостимуляторов и т. и.).
Разу-меется, реакция на воздействие начинает формироваться немедленно.
Вначале развивает-ся первая быстрая, нервная фаза реакции. При этом
изменяются разные показатели, в том числе и лейкограмма. Так, например,
после активационных воздействий характерное для реакции
активации относительное число лимфоцитов, отмечающееся через 6 и 24 ч,
об-наружено уже через 5—15 мин (Бутов М. А., 1984, 1985; Бутов М.
А., Ногаллер А. М., 1986). Затем отмечаются волнообразные изменения,
включается медленное гуморальное звено формирования реакции и к 6 ч
после воздействия реакция организма околосуточно-го ритма уже
сформирована, а колебания наблюдаются в пределах данной реакции.
Нару-шение такой зависимости может
быть лишь при
дополнительном появлении сверх-сильного раздражителя.
В ЦНС в стадии первичной активации преобладает умеренное
физиологическое возбуждение. На уровне поведенческих реакций это
проявилось в умеренном повышении общей двигательной активности (Гаркави
Л. X., 1968; Шихлярова А. И., 1975; Марьяновская Г. Я., Зигун Л. Н.,
1985). Возбудимость нервных структур гипоталамуса повышена на
20—35% по сравнению с исходной (см. рис. 16), в ЭЭГ происходит
сдвиг спектра в сторо-ну быстрых волн с уменьшением амплитуды (Квакина
Е. В., 1972; Котляревская Е. С, 1974). Вероятно, с этим связан тот
факт, что реакцию активации удается легко вызывать с помощью
переменного магнитного ноля (ПеМП), так как в отличие от постоянного
ПеМП может вызывать в мозгу преобладание процессов возбуждения (Квакина
Е. Б., Уколова М. А., 1969, 1971; Квакина Е. Б., 1972). С помощью ПеМП
реакцию активации получали в эксперименте и другие авторы (Макеев В.
Б., Темурьянц Н. А., 1982; Сидякин В. Г., Те-мурьянц Н. А.. Евстафьева
Я. В., 1986).
Исследование сверхмедленных колебаний потенциалов головного мозга у
людей показало, что амплитуда колебаний при развитии реакции активации
больше, чем при стрессе, но меньше, чем при реакции тренировки
(15—40 мВ) и преобладают более длин-ные волны (период 1,5-2 ч).
Были получены четкие различия по показателю асимметрии
?-волны в ЭЭГ — ?, отражающему регулярность цикла
«возбуждение — торможение» (Генкин А. А., 1965). Если
стресс характеризовался кривыми ?, с нерегулярными
коле-баниями, то при реакциях тренировки и активации отмечались более
регулярные колеба-ния ?, причем реакцию активации отличали колебания ?
несколько большей частоты и амплитуды (Коробейникова Е.
П., 1978).
Так как
чувствительность в стадии первичной
активации повышается, то для получения второй стадии
реакции дозу необходимо немного
уменьшить. Однако это процесс
волнообразный, и после
45
уменьшения дозы в дальнейшем ее приходится несколько увеличивать, затем
— опять уменьшать и т. д. Вторая стадия реакции активации
развивается при систематическом по-вторении раздражений средней силы.
Она названа нами стадией «стойкой активации», так как при
ней стойко держится активация защитных систем организма. Тимус также
увели-чен, соотношение форменных элементов белой крови остается таким
же. Гормональная характеристика этой стадии существенно не отличается
от стадии первичной активации. Содержание 17-ОКС и 11-ОКС, кортизола
претерпевает колебания от нормы до величин, превышающих норму примерно
в 1,5 раза. Такие же изменения происходят с содержанием калия в крови,
содержание натрия все время повышено (на 25—40%). Содержание БСЙ
повышено в первые 2 — 3 недели воздействия, а затем существенно
не отличается от кон-троля, а БЭЙ, тироксина и трийодтиронина все время
несколько повышено. Наличие вы-сокой функциональной активности в этой
стадии подтверждается и морфофункциональ-ными изменениями щитовидной
железы: увеличена высота тиреоидного эпителия (12,1 ±0,3 мкм по
сравнению с 8,0 ±0,7 мкм контроля), а также размер его ядер (6,1
±0,1 мкм но сравнению с 5,4±0,1 мкм контроля). Активность
половых желез также умеренно повыше-на; несколько увеличена
продолжительность активных фаз цикла. Повышена гонадотроп-ная
активность гипофиза. В гипофизе увеличивается число гонадотрофов
(11±2,2 по сравнению с 5,2±0,6 контроля) и тиреотрофов
(9,6±4,2, в контроле — 3,2±0,9), т. е. изменение
клеточного состава гипофиза также говорит о повышении активности
щитовидной и по-ловых желез. В то же время число ацидофилов в гипофизе
не увеличено (25,8±2,8, в контроле —30,6±3,0—
разница несущественна).
Показатели метаболизма в этой стадии претерпевают различные изменения.
Со-держание аммиака остается умеренно повышенным — в 1,5—2
раза, содержание глюта-мина и глютаминовой кислоты умеренно снижено
— на 20—30%. Показатели энергетиче-ского обмена мозга также
меняются фазно. Через неделю тканевое дыхание повышается (на 25%),
накопление АТФ и КФ, а также эстерификация неорганического фосфора
сни-жаются (соответственно на 35 и 43%). Анаэробный гликолиз снижается
до уровня контро-ля, а аэробный — до уровня ниже нормы (на 37%).
Через 4 недели тканевое дыхание, на-копление АТФ и КФ возвращаются к
уровню контроля. Эстерификация неорганического фосфора немного снижена
(на 20%), анаэробный и особенно аэробный гликолиз вновь по-вышается
(Гаркави Л. X. и др., 1968).
Полярографическое исследование энергетического обмена на уровне
митохондрии показало изменения, сходные с 1-й стадией, но с
возрастанием фонда ЭЯК, основного субстрата окисления (Гаркави Л. X.,
Квакина Е. Б., Котляревская Е. С, Барсукова Л. П.,
46
Марьяновская Г. Я., 1987). Содержание РНК в различных органах также
претерпевает фазные изменения: первоначальное повышение сменяется через
2 недели снижением (на 15—35%), а затем вновь происходит
повышение (на 15—25%). В течение первых двух не-дель. отмечается
умеренное повышение белка и белковых фракции (на 35—40%), а через
3—4 недели воздействий — такое же умеренное снижение.
У людей в сыворотке крови при длительном поддержании реакции
активации со-держание общего белка — на уровне верхней границы
нормы, альбумино-глобулиновый коэффициент выше, чем при стрессе, но
несколько ниже, чем в контроле, в пределах ниж-ней половины зоны нормы.
Такое соотношение связано с увеличением гамма-глобулинов. При стрессе,
напротив, количество альбуминов значительно уменьшено, а содержание
всех фракций глобулинов резко увеличено.
Таким образом, относительное постоянство нейрогормональных отношений в
ста-дии стойкой активации поддерживается фазными изменениями некоторых
показателей метаболизма.
При развитии реакции активации отмечается физиологическая активация
элемен-тов ретикулярной формации межуточного мозга без каких-либо
деструктивных изменений (Гаркави Л. X., 1969; Квакина Е. Б., Шибкова С.
А., Исаджанова С. X., 1973). Отмечены признаки повышения функциональной
активности нейронов: гиперхроматоз цитоплазмы при окрашивании на РНК и
тигроид, повышение аргентофильности, гипертрофия нейро-нов (рис. 27), в
отличие от гипохроматоза нервных клеток при стрессе (рис. 28). Имеются
данные, что при развитии стресса (в том числе, при росте опухоли) в
ретикулярной фор-мации межуточного мозга отмечаются деструк тивные
изменения: распад ретикулярных и нервных клеток, распад
аксосоматических синапсов и т. д. (Чаусов В. И. и др., 1967).
Как известно, большое значение в обеспечении функционально" перестройки
ЦНС придается метаболизму нейроглии и глионейрональным соотношениям. М.
М. Александ-ровская отмечает наличие параллелизма между величиной
функциональных сдвигов и ха-рактером изменений нейроглии. Слабые
воздействия — токсические агенты в малых до-зах, малые дозы
облучения вызывают активацию нейроглии в коре головного мозга, в то
время как тяжелая интоксикация приводит к угнетению нейроглии,
сопровождающемуся дистрофическими изменениями и гибелью нейронов. М.М.
Александровская (1968) при-ходит к выводу, что «реакция нейроглии
и глионейрональные соотношения могут быть тонкими индикатором и
изменений функционального состояния ЦНС». Роль глии в реак-ции
ЦНС на МП неоднократно подчеркивалась Ю. А. Холодовым (1975 1979).
Нами отмечалось повышение функциональной активности нейро-
47
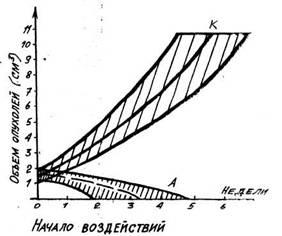
Рис. 27. Мозг крыс. Реакция активации: ретикулярное ядро, гиперхромные нервные клетки. Окр. по Браше. Ув. 100Х 6,3
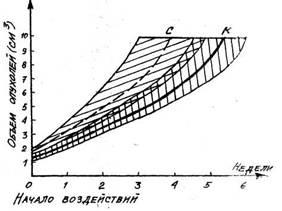
Рис. 28. Мозг крыс. Реакция
стресс: ретикулярное ядро,
гипохромные нервные клетки. Окр. по Браше. Ув. 100X6,3
глиальных элементов: обогащение клеток гликогеном и белками,
пролиферация клеточ-ных элементов и сателлитов серого вещества мозга.
Наиболее выраженные изменения наблюдались в микроглиаль-ных клетках:
число их отростков увеличено, некоторые из них были утолщены и
выглядели фрагментирован-ными. Особенно наглядным было то, что на
перикарионах нейронов гипоталамуса распо-лагались группы ядер
нейроглиальных клеток (рис. 29), в то время как в гипоталамусе
контрольных крыс ядра глиальных клеток лежали, как правило, между
нейронами и, ред-ко, на поверхности перикариона. Факт нахождения
глиальных клеток на перикарионах нейронов Свидетельствует об усилении
глионейрональных контактов, так необходимых при повышенной
функциональной активности нейронов. Кроме этого, повышалась
функ-циональная активность синапсов синапсы увеличены, аргентофильность
их повышена (рис. 30).
Возбудимость (по показателям порогов раздражения) в стадии стойкой
активации остается умеренно повышенной, в ЭЭГ так же, как и в стадии
первичной активации, пре-обладают более высоко-
48
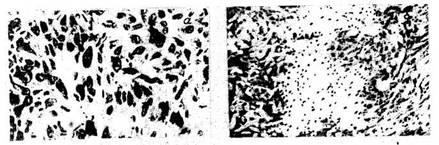
Рис. 29. Мозг крыс. Реакция активации: латеральное гипоталамическое
ядро, расположение ядер сателлитов на поверх-ности перикарионов крупных
нейронов. Окр. по Эйнарсону. Ув. 100X6,3
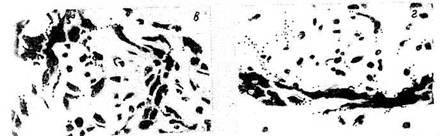
Рис. 30. Мозг крыс. Реакция активации: увеличение размеров синапсов в
мотосенсорной области коры головного мозга и усиление импрегнации.
Импрегнация серебром по методу Кахала-Фаворского. Ув. 40Х 7
49
частотные колебания, чем при реакции тренировки.
В результате исследований было найдено, что в реакции активации можно
выде-лить 2 зоны, отличающиеся друг от друга: спокойной активации и
повышенной актива-ции. Как говорит само название, зона спокойной
активации (ЗСА) лежит ближе к реакции тренировки, а зона повышенной
активации (ЗПА) — к стрессу. Степень активации защит-ных систем
организма при ЗПА больше, чем при ЗСА. Дальнейшие исследования
показа-ли, что различия настолько велики, | что можно назвать зону
спокойной активации — ре-акцией спокойной активации
(РСА), а зону повышенной активации — реакцией I повы-шенной
активации (РПА). В тимусе у крыс в РСА при значительном увеличении
фигур митоза в дольках отмечено равномерное расширение корковой
части или внедрение тя-жей корковой зоны - в
мозговую. В тимусе крыс в
РПА наблюдается разной степени выраженности
гипертрофия мозговой части. В лимфатических узлах при РСА отмечена
выраженная плазматизация лимфоидных элементов, а при РПА, в большинстве
случаев — гиперплазия клеток ретикулоэндотелия.
Кроме того,
усиление дифференцировки
пиронинофильных лимфоцитов в зрелые формы, характерное и для
фолликулов, у крыс в РПА выражено
значительно больше. Такие
участки занимают большую часть лимфоузлов.
В селезенке крыс в РСА число фолликулов в 2—4 раза больше, чем в
реакции тре-нировки. Изменяется соотношение размеров герминативных
центров и пояса короны ма-лых лимфоцитов в пользу последних. В РПА в
большинстве фолликулов стирается грань поясов при полной или почти
полной дифференцировке бластов герминативных центров в зрелые формы
(Мулатова А. К., 1983).
Определены параметры реакций спокойной и повышенной активации по
составу белой крови. При РСА число лимфоцитов — в пределах
верхней половины зоны нормы (28—33%), число сегментоядерных
нейтрофилов — в пределах нижней половины зоны нормы, число
лейкоцитов, эозинофилов, палочкоядерных нейтрофилов и моноцитов —
в пределах нормы. При РПА число лимфоцитов несколько выше нормы, более
33%, до 40—45% (индивидуально), число сегментоядерных нейтрофилов
— ниже нормы, число лейкоцитов, эозинофилов, палочкоядерных
нейтрофилов и моноцитов — в пределах нормы.
Удалось выявить заметное отличие между РСА и РПА и по гормональным
показа-телям. Так, у людей при РСА содержание корти зола в крови
составляет 197 ± 67,8 нмоль/л, при РПА — 301 ± 74,1
нмоль/л; при РСА содержание в крови тироксина —129 ± 9,4
нмоль/л, при РПА —154 ± 7,9 нмоль/л. Описанные ранее для
стадии первичной акти-вации морфологические признаки повышения
функциональной активности щитовидной железы также более ярко выражены в
РПА.
50
При сравнении гормональных изменений необходимо
подчерк нуть значение не столько абсолютных количеств содержания
гормонов, сколько гормональных отношений. Так, по содержанию глю
кокортикоидов реакция тренировки мало отличается от повы-шенной
активации, а отношение глюкокортикоиды — гормоны щитовидной
железы отли-чается в 2—3 раза. Иначе говоря, при сходном уровне
глюкокортикоидов функциональная активность щитовидной железы при
повышенной активации значительно выше.
Есть отличия и по изменениям свертывающей системы крови. При РСА
изменения такие же, как описанные для стадии первичной активации. При
РПА развивается умерен-ная гипокоагуляция. Гак, увеличивается время
реакции «R» (11,8±1,2 мм по сравнению с
6,8±О,6 мм), противосвертывающая система (R+ K) активируется.
При спокойной активации скорости окисления субстратов и окислительного
фос-форилирования (но сравнению с реакцией тренировки)
возрастают, а время
фосфори-лирования уменьшается. Наряду со
снижением фонда ЭЯК увеличивается ее воспроиз-водство. При повышенной
активации, кроме того, выявляется неглубокое ЩО.
У здоровых РПА можно поддерживать длительное время. РПА—-наиболее
стойкая реакция у здоровых. РСА — не столь стойкая реакция. У
больных, особенно тяжело боль-ных, РПА, напротив, поддерживать трудно,
так как если своевременно не уменьшить дозу (силу) воздействия, то РПА
легко переходит в стресс, а иногда — в резкую переактивацию с
чрезмерно увеличенным числом лимфоцитов и отклонением общего числа
лейкоцитов либо в сторону лейкопении (чаще), либо в сторону
лейкоцитоза.
Реакция активации, как спокойная, так
и повышенная, легко вызывается с
помощью умеренных раздражений
положительных эмоциональных зон мозга, тогда как реакция тренировки
— при слабом раздражении, а стресс — сильном раздражении
от-рицательных эмоциогенных зон. По-видимому, это связано с разной
продукцией эндор-финов при этих реакциях (Гаркави Л. X., Квакина Е. В.,
За харова Н. П., Шихлярова А. И., Квакина Т. С, Коробейникова Е. II.,
Мамцева Л. И., 1982; Гаркави Л. X., Евстратова О. Ф., Коробейни нова Е.
П., 1983; Гаркави Л. X., Квакина Е. В., Евстратова О. Ф., 1983).
Таким образом, несмотря на существенные отличия реакций спокойной и
повы-шенной активации, комплекс изменений, характерный для них, может
быть назван анти-стрессорным.
Резистентность организма при развитии реакции активации
Как уже говорилось, резистентность организма повышается в стадии
резистентно-сти стресса и в различных стадиях реакции тренировки.
Однако характер повышения ре-зистентности и цена резистентности при
каждой стадии этих реакций различны.
51
При развитии реакции активации как спокойной, так и повышенной, уже в
стадии первич-ной активации происходит истинное повышение активной
резистентности организма: не за счет развития торможения и снижения
чувствительности, а за счет истинного подъема активности защитных
систем организма. Повышение активной резистентности сохраняет-ся и
приобретает устойчивость, стабильность в стадии стойкой активации,
особенно по-вышенной (рис. 31). Резистентность при реакции активации не
только быстро

Рис. 31. Схема изменения резистентности организма: а
— при стрессе, б — при реакции активации
повышается и стойко держится при повторении активирующих воздействий,
но и, в отли-чие от реакции тренировки, остается повышенной в течение
некоторого времени и после прекращения воздействий. Длительность
последействия широко варьирует в зависимости от состояния здоровья и
возраста — от 1-2 недель до полугода. Необходимо подчеркнуть, что
такое последействие наблюдается лишь в стадии стойкой активации, т. е.
после дли-тельного, систематического повторения активирующих
воздействий.
Энергетические траты при реакции активации хотя и выше, чем при реакции
тре-нировки, но напряжение, как при стрессе, не развивается. Кроме
того, хорошая уравнове-шенность различных видов метаболизма приводит к
тому, что не только не преобладают явления распада, но даже, наоборот,
происходит накопление строительного материала — аминокислот,
нуклеиновых кислот, белков.
Реакция активации, особенно повышенная — анаболическая реакция.
По этому по-казателю, так же, как и по многим другим, РПА
напоминает «физиологический
стресс» И. А.
Аршавского (1982, 1986).
Преобладание минералокортикоидов — прововоспалительных гормонов
— и весь комплекс изменений в нейроэндокринной системе при
реакции активации говорят об уме-ренном повышении прововоспалительного
потенциала организма.
Как и при реакции тренировки, повышение резистентности при
52
реакции активации исследовалось но отношению к повреждающему действию
токсиче-ских противоопухолевых препаратов и облучению в терапевтических
дозах, а также к опухолевому процессу.
Электрическое раздражение гипоталамуса (средней силы), переменное
магнитное поле и адреналин в средних дозах уже при однократном
воздействии (до облучения) уменьшали лейкопению и летальность. При
общем облучении крыс в.дозе 6 Гр через 6 су-ток в опытной группе число
лейкоцитов было 3575±376, а в контрольной — 1800 ±
34 (р<0,05). В опытах на выживаемость с облучением в дозе 7 Гр в
контрольной группе пало 40% крыс, в то время как в опытной лишь 10%.
Многократное повторение тех же воздей-ствий в средних дозах
поддерживало состояние высокой резистентности в течение дли-тельного
времени (Гаркави Л. X., Квакина Е. В., Тарасьянц Ф. М., Уколова М. А.,
1976; Гаркави Л. X., Квакина Е. Б, Мулатова А. К., Тарасьянц Ф.
М., 1980). При локальном об-лучении крыс в дозе 20 Гр наблюдалось
уменьшение послелучевой лейкопении и затем более полная нормализации
белой крови в подопытной группе по сравнению с контроль-ной. Через 1 и
2 недели число лейкоцитов в контрольной группе снизилось
соответствен-но на 7505±870 и 4910±1120, а в подопытной
на 4750±450 и 2460±750 (Комурджиев X. А. и др., 1968;
Квакина Е. В., 1972). Такое же защитное действие отмечалось при
введении крысам сарколизина — по 2 мг/кг, суммарная доза 42
мг/кг, и циклофосфана — по 7 мг, суммарная доза 147 мг/кг
(Захарюта Ф. М., 1983). Об этом же свидетельствуют, например, изменения
в лимфоузлах (рис. 32).
При длительном введении противоопухолевых антибиотиков —
оливомицина (по 0,03 мг через день, всего 10 раз) и актиномицина (по
2—3 мг через день, всего 10 раз) и одновременного применения
ионизирующего облучения (в дозе 2,5 Гр через день, также 10 раз)
— удалось защитить организм от повреждающего действия этих
факторов. По-скольку облучение в этих опытах применялось в дозе, не
вызывающей лейкопении, а при-менявшиеся препараты не влияют
избирательно на кроветворение, то о резистентности животных судили по
общему состоянию и весу, массе тимуса, морфологии и гистохимии коры
надпочечников, играющих важную роль в адаптации организма к
повреждающим факторам. Животные после облучения, введения оливомицина и
актиномицина были вя-лыми, малоподвижными, с редкой тусклой желтой
шерстью. После введения оливомицина, кроме того, отмечались носовые
кровотечения, а актиномицина — диаррея. При со-вместном
применении этих препаратов с об лучением такие явления были особенно
вы-ражены. У крыс наблюдалась потеря веса (16 ± 2,8
г—актиномицин; 21 ± 3,1 г —актино-мицин +
облучение; 18 ± 2,2 г — оливомицин; 31 ± 4,3 г
— облучение; 32 ± 3,6 г — облу-чение + оливомицин).
Масса тимуса у крыс с облучением и введением оливомицина и
ак-тиномицина была сни-
53

Рис. 32. Лимфатический узел крысы: а —
реакция стресс после
введений сарколизина. Резкая гипо-плазия лимфоидных
элементов. Образование кист; б —
реакция активации при введении
сарколизина в той же дозе на фоне
введения адреналина. Кисты отсутствуют. Гиперплазия
клеток ретикулярной
ткани и лимфоидных элементов. Окр.
по Браше в модификации А. А.
Симаковой. Ув. 8X7
жена (до 70±5,7 мг). Животные, у которых поддерживалась реакция
активации, напротив, были подвижны, носовых кровотечений и диарреи не
отмечалось, шерсть была густой, белой, блестящей, крысы не теряли в
весе, тимус не уменьшался (160±4,3 мг).
При облучении и введении противоопухолевых препаратов отмечались
деструктивные изменения во всех зонах коры надпочечников,
особенно клубочковой и
сетчатой (резко выраженная
зернистая и вакуольная дистрофия, отек межклеточных про-странств).
Гистохимические исследования
содержания липидов, ДНК,
кислых и нейтральных мукополисахаридов также
свидетельствовали о нарушении функции
коры надпочечников, особенно
клубочковой зоны, ответственной за секрецию
минерало-кортикоидов. У крыс, подвергавшихся систематическим
воздействиям, вызывающим раз-витие реакций активации,
деструктивные изменения в надпочечниках были
значитель-но менее выражены. Содержание липидов в клубочковой
и сетчатой зонах было
не-значительным, липиды определялись,
54
главным образом, в пучковой зоне. Количество ДНК было увеличено в
клетках клубочко-вой зоны. Все эти изменения свидетельствовали о
повышении функциональной активно-сти преимущественно клубочковой зоны
(Гаркави Л. X., Колесникова В. Д., Андреева Л. А., 1971; Огородникова
Л. С, Гаркави Л. X., Андреева Л. А., 1973). Таким образом, разви-тие
реакции активации повышало резистентность организма к повреждающему
действию лучевой и химиотерапии, вызывающим стресс.
Особенно удивительным был тот факт, что повышение резистентности при
под-держании реакции активации оказалось достаточным, чтобы вызвать
полную регрессию уже развившихся экспериментальных опухолей как
перевивных, так и индуцированных. Вместе с тем уже не единичны случаи
самопроизвольного торможения и рассасывания опухолей (Швембергер И. Н.,
1987). Это, а также наличие низкораковых линий животных и
животных-нуллеров, у которых опухоли не возникают, позволяет
предположить, что при каком-то функциональном состоянии организм
становится резистентным по отноше-нию к опухоли. О значении
регуляторных и защитных систем организма в формировании
противоопухолевой резистентности свидетельствуют работы Р. Е. Кавецкого
(1962, 1973), И. П. Терещенко (1967, 1976, 1984), К. II. Валицкого и И.
Г. Векслера (1975, 1983), В. В. Винницкого (1983) и др. Ранее уже
говорилось о состоянии организма, вызываемого адап-тогенами, описанном
Н. В. Лазаревым — состоянии неспецифически повышенной
сопро-тивляемости (СНПС). По мнению В. Г. Нашинского и К. В. Яременко
(1983), интенсивно развивающих онкологическую фармакотерапию с помощью
средств природного проис-хождения, СНПС имеет много общего с реакциями
активации и тренировки, так как при нем происходит коррекция нарушенных
физиологических функций, усиление активности защитных систем организма
и ухудшение тем самым условий существования опухолевых клеток.
Функциональное состояние организма, при котором повышена его
противоопухо-левая резистентность, развивается при общей не
специфической адаптационной реакции активации. Многочисленные серии
опытов на животных (более 10 тыс.) подтвердили этот факт (Гаркави Л.
X., 1964, 1968, 1969; Уколова М. А., Квакина Е. В., 1970, 1971;
Бордюш-ков Ю. Н., 1971; Уколова М. А., Гаркави Л. X,, Квакина Е. В.,
1973; Гаркави Л. X., Уколо-ва М. А., Квакина Е. Б., 1973: Марьяновская
Г. Я., 1974, Котляревская Е. С, 1974; Шихля-ро-ва А. И., 1975; Гаркави
Л. X., Квакина Е. Б., Мулатова А. К., Ших-лярова А. И., 1980; Шихлярова
А. И., 1985, 1986; Марьяновская Г. Я.. Барсукова Л. П., Котляревская Е,
С, Квакина Е. Б., Гаркави Л, X.,. 1986).
Для развития и поддержания реакции активации с целью повышения противоопу-холевой резистентности применяли те же воздей-
55
ствия: электрическое раздражение гипоталамуса, низкочастотные
переменные магнитные поля, адреналин, карбахолин, холинэстеразу,
эфедрин, этимизол, мелипрамин, мумие, элеутерококк колючий, корень
левзеи, прополис. Использовались различные штаммы экспериментальных
опухолей крыс, на которых обычно изучалась противоопухолевая
эффек-тивность препаратов: саркома-45, саркома-М-1, гепатома,
асцитная опухоль яичников, опухоли яичника по Бискиндам, карцинома
Герена, рак молочной железы крыс, генерация сарком, получаемых при
последовательных перевивках индуцированных сарком, и сарко-мы
первично-индуцированные химическими канцерогенами:
3—4-бензапиреном и метил-холантреном. Независимо от вида опухоли,
противоопухолевый эффект наблюдался при развитии стойкой активации.
Воздействия начинали после того, как опухоли достигали величины, после
которой они самопроизвольно не рассасываются (более 1 см в диаметре), а
в опытах с влиянием на «большие» опухоли (больших сроков
развития) — по достижении 11,5 см в диаметре. Воздействия
повторялись ежедневно до полного рассасывания опухолей, в отдельных
се-риях опытов процент рассасывания колебался от 30 до 100%, у животных
без полной рег-рессии опухоли примерно в половине случаев отмечалось
торможение роста по сравне-нию с контролем. Эффективность воздействий в
какой-то мере зависит от характера опу-холей — преимущественно от
присущего им темпа роста. Но, главным образом, эффект зависел от того,
удавалось ли вызвать и стойко поддерживать реакцию активации. Во всех
случаях, когда это удавалось, отмечался противоопухолевый эффект вплоть
до полной регрессии опухоли (рис. 33). У тех животных, у которых
стойкой активации получить не удалось, опухоли продолжали расти. В
опытах с большими опухолями выяснилось, что наиболее эффективной
является реакция повышенной активации (РПА). Рассасывание больших
опухолей происходило тогда, когда удавалось вызвать и стойко
поддерживать РПА. Таким образом, в тех случаях, когда удавалось вызвать
и поддерживать стойкую ак-тивацию, особенно РПА, всегда получали
противоопухолевый эффект. При специальном вызове и поддержании стресса
(большие дозы) рост опухолей усиливался (рис. 34).
Характер изменений в нейроэндокринной системе напоминал реакцию
активации у животных без опухоли, но имел и следы опухолевого стресса.
В случаях, когда противо-опухолевого эффекта получить не удавалось,
изменения были характерными для стресса. При рассасывании опухолей
содержание в крови людей 17-ОКС и 11-ОКС в крови крыс было немного выше
нормы (на 40—60%), содержание калия существенно не отличалось от
нормы, содержание натрия немного повышено (на 25—30%), содержание
БСЙ повы-шено (на 60 — 70%), БЭЙ также повышено (на
50—60%). В то же время при росте опухо-лей
56
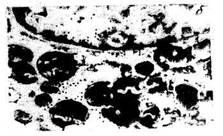
Рис. 33. Рост экспериментальных
опухолей: К—без воздействия, А
— при активационных воздей-ствиях на организм

Рис. 34. Рост экспериментальных опухолей: К — без воздействия, С — при стрессорных воздействиях на организм
(в период бурного роста) содержание 17-ОКС и 11-ОКС резко повышено
— в 3—3,5 раза, содержание калия повышено (на
65—70%), а натрия, напротив, несколько снижено (на 20—25%),
содержание БСЙ и БЭЙ снижено (соответственно на 30—40% и
40—50%). В надпочечниках при рассасывании опухолей отмечались
признаки повышенной активности клубочковой зоны (высота зоны 90,2
± ±3,9 мкм, как по сравнению с надпочечниками крыс без
опухолей 78 ± 10 мкм, так и, особенно, с надпочечниками крыс с
растущими опухолями 66,6 ±3,2 мкм); количество липидов и
аскорбиновой кислоты уменьшалось во всех зонах, особенно в клубочковой.
В щитовидной железе наблюдалось увеличение вы-соты тиреоидного эпителия
(12,9±0,2 мк, в контроле —9,5±0,15 мк) и
увеличение раз-
57
меров его ядер (6,03 ± 0,13 мк, в контроле —
5,6±0,1 мк). Вес тимуса при рассасывании перевивных опухолей
приближается к норме. При гистологическом исследовании в кор-ковом слое
тимуса и лимфатических узлах отмечается гиперплазия лимфоидных
элемен-тов. При росте перевивных опухолей масса тимуса снижена на
55—65%, а при росте ин-дуцированных опухолей составляет лишь
8—10% нормы. Иными словами, при рассасы-вании опухоли тимус
увеличивается в 5 —11 раз (Гаркави Л. X., 1968, 1969; Квакина Е.
Б., Николаева В. И., Уколова М. А., 1971; Квакина Е. Б., 1972).
Морфологический состав белой крови при рассасывании опухоли у крыс
соответ-ствует характеристике реакции активации. Число лейкоцитов
— в пределах нормы (6,0 • 109—20,0 -109), число
эозинофилов — в пределах нормы, палочкоядерные — в пределах
нормы, сегментоядерные — в пределах нижней половины зоны нормы
(меньше 30%), лимфоциты — в пределах верхней половины зоны
нормы (60—80%), моноциты — в пре-делах нормы.
Активность дегидрогеназ лимфоцитов (СДГ и ЛДГ) при рассасывании
опухолей существенно не отличалась от исходных величин животных без
опухолей. Существенно, почти вдвое, повышалась активность ?-ГФДГ
цитоплазматической, являющаяся, по неко-торым данным, маркером
иммунного ответа (Зарецкая Ю. М., 1983).
Изменения различных показателей метаболизма при рассасывании опухолей
также были близки к наблюдаемым при реакции активации. Содержание
аммиака в мозгу было умеренно повышено (в 1,5 раза), глютамин и
глютаминовая кислота — на нижней границе нормы. Тканевое дыхание
мозга несколько повышено (на 25 — 30%), накопление АТФ+КФ также
повышено (45—55%). Эстерификация неорганического фосфора и
ана-эробный гликолиз существенно не отличались от нормы, аэробный
гликолиз был повы-шен на 80—90% (Гаркави Л. X., Чернявская Г. Я.,
1970; Уколова М. А., Квакина Е. Б., Чернявская Г. Я., 1969).
Полярографическое исследование энергетического обмена на уровне
митохондрий показало тенденцию к нормализации энергетического обмена по
по-казателям скоростей окисления ЯК, фонда ЭЯК, воспроизводства ЯК и
уменьшению глу-бины щавелево-уксусного ограничения (Марьянов-ская Г.
Я., Барсукова Л. П., Котлярев-ская Е. С, Квакина Е. Б., Гаркави Л. X.,
1986). Имеется тенденция к нормализации содер-жания общего белка и
особенно белковых фракций тканей и сыворотки крови.
При росте опухолей происходят изменения, близкие к действию сильного
раздра-жения. Как уже говорилось, вес тимуса при росте опухолей резко
уменьшен. Картина бе-лой крови типична для стресса не острого, а
хронического — длительно текущей при опухолевой болезни стадии
истощения. Отличия от острого стресса таковы: не обязатель-но наличие
лейкоцитоза — может быть и нормальное число
58
лейкоцитов и даже лейкопения; отсутствие эозинофилов так же не
обязательно — может быть и нормальное число эозинофилов и даже
эозинофилия. Эозинофилия в сочетании с лимфопенией — плохой
прогностический признак, так как свидетельствует об истощения
глюкокортикоидной функции коры надпочечников (Мацанов .А. К.. 1971).
Изменения в эндокринной системе типичны для стресса, за исключением
того, что в части случаев при больших опухолях секреция
глюкокортикоидов была не повышена, а даже снижен:), что согласуется с
данными литературы (Самунджан Е. М., 1975) Изменения метаболизма при
росте опухолей также были типичны для хронического стресса. Содержание
аммиака зна-чительно повышено (в 3 раза), содержание глютамина
повышено (на 20—25%), содержание глютаминовой кислоты
снижено (на 50—55%). При росте опухолей тканевое дыха-ние и
накопление АТФ и КФ несколько снижены (на 15—20%), снижены
эстерификация неорганического фосфора (на 30—35%) и анаэробный
гликолиз (на 25-30%), а аэробный — резко повышен (в 2—3
раза). При росте опухоли в печени отмечались выраженные признаки
низкоэнергетического сдвига. Интенсивный рост опухоли сопровождался
либо снижением, либо резким повышением скоростей дыхания, существенным
увели чением времени фосфорилирования и снижением реакции на повторную
добавку АДФ, вплоть до ее полного отсутствия. Кроме того, отмечались
резкий дефицит ЯК и углубление степени щавелево-уксусного ограничения
активности СДГ. Наблюдалось резкое снижение альбу-мино-глобулинового
коэффициента, общего белка и белковых фракций в тканях и сыво-ротке
крови. Содержание РНК в тканях также значительно снижено.
Исследование морфологических и гистологических изменений в опухоли и
окру-жающих тканях в динамике показало, что процесс рассасывания идет,
главным образом, с периферии. На периферии опухоли происходит
разрастание волокнистой соединительной ткани и врастание ее из
окружающих тканей в опухоль. Отмечается лимфоцитарная ин-фильтрация
опухоли. Опухолевые клетки подвергаются некротическим и
некробиотиче-ским изменениям с последующим лизисом, содержание в них
мукополисахаридов и нук-леиновых кислот постепенно уменьшается.
Аргирофильные волокна вначале иг определя-лись, а затем выявлялись в
виде крупнопетлистой сети. Число митозов опухолевых клеток резко
уменьшалось. На конечном этапе рассасывания вместо бывшей опухоли можно
бы-ло наблюдать лишь участок, состоящий из рыхлой волокнистой
соединительной ткани и небольшого количества лимфоцитов. Опухолевые
клетки не определялись. Механизм рас-сасывания был единым и в случае
перевиваемых, и в случае первично-индуцированных опухолей (рис. 35).
Это местное проявление общей реакции свидетельствует об активации
тимико-лимфатической системы и системы соединительной ткани.
59
Рис. 35. Саркома-45:
а — через 1,5 недели (10 дней) после перевивки до начала
воздействий. Интенсивный рост
веретенокле-точной саркомы. Окр. галлоцианином по
Эйнарсону в модификации де Бера и Сарна-кера.
Ув. 40Х 10; б — через 18 дней после
перевивки и через 7 дней воздействий. Участки регрессии опухоли. Окр.
та же. Ув. 16Х10 и 40Х10;
в — через 25 дней после перевивки и через 14 дней
воздействий. Почти полная регрессия
опухоли с сохранившимися тяжами опухолевых
клеток. Окр. та же. Ув. 40 X 10;
г — через 30 дней после перевивки и через 19 дней воздействий. На
месте бывшей опухоли фибробластиче-ский рост.
Отдельные мелкие пласты
опухолевых клеток. Окр. та же. Ув.
40Х 10;
д — пролиферация эпителия в
сохранившихся придатках кожи при
замещении опухоли соединительной тканью.
Окр. та же. Ув. 16X10
60
Исследования ультра- и микроструктуры опухолевых клеток в процессе
рассасыва-ния опухоли характеризовались изменениями эндоплазматического
ретикулума, набуха-нием митохондрий и расплавлением их внутренних
мембран и крист (рис. 36). С помощью микроспектрального люминесцентного
метода с использованием акридинового оранжево-го получены данные о
значительном снижении уровня синтетических процессов в цито-плазме и
ядрах опухолевых клеток (Шихлярова А. И., 1985).
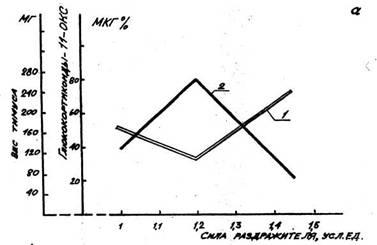
Рис. 36.
Саркома-45:
а — рост опухоли в контроле.
Ядро занимает большую часть клетки, хроматин в виде
грубых глыбок неравномерно
распределен по ядру. В цитоплазме
опухолевой клетки система мембран
эндоплазма-тического ретикулума с большим
количеством РНП-гранул. Электр.
микр. Ув. X 10000;
б — регрессия опухоли.
Ядерная оболочка местами выделяется
нечетко. Хроматин разрежен, имеются поля,
лишенные хроматина.
Гранулярный эндоплазматический ретикулум
сохранен в виде единичных фрагментов с небольшим количеством РНП-гранул
на мембранах. Увеличено число лизосом. Электр.
микр. Ув. X 14000;
в —регрессия
опухоли. Разрежение ядерного хроматина,
разрушение ядерной оболочки. Электр. микр. Ув. X 7000;
г — регрессия опухоли. Увеличение размеров
митохондрий, расплавление их
мембран и крист. Электр. микр. Ув. X10000
61
Необходимо специально остановиться на роли лимфоцитов в повышении
неспеци-фической, в том числе противоопухолевой резистентности при
реакции активации. При описании реакции активации говорилось, что
повышение процентного содержания лим-фоцитов (до уровня верхней
половины зоны нормы и выше) периферической крови явля-ется неотъемлемым
признаком этой реакции как при отсутствии опухоли, так и при ее
на-личии (если удается вызвать реакцию активации). Информативность
этого показателя для оценки противоопухолевой резистентности отмечается
и в других работах (Волегов А. И., 1987). В тимусе и лимфатических
узлах наблюдается гиперплазия лимфоидных элемен-тов. Рассасывание
опухоли происходит при внедрении лимфоцитов в опухолевую ткань,
инфильтрации опухоли лимфоцитами. И вся эта картина повышения
активности лимфо-цитов, четко выявляющаяся при изучении даже только
количественных показателей, раз-вертывается стандартно в ответ на
действие самых различных неспецифических (в отно-шении иммунитета)
раздражителей средней силы. В настоящее время научная литература
насыщена данными о роли различных популяций лимфоцитов в иммунных
реакциях, в том числе, и в формировании противоопухолевого иммунитета.
Однако этот вопрос изу-чается, как правило, без связи с общими
неспецифическими адаптационными реакциями организма. Между тем иммунная
система, хотя и очень важная, но всего лишь подсистема организма,
безусловно подчиняющаяся количественно-качественным закономерностям
развития неспецифических адаптационных реакций целого организма.
Сопоставление ти-па адаптационной реакции, например, с показателями
клеточного иммунитета у больных раком легкого, показало четкую
корреляцию между характером реакции и количеством Т- и В-лимфоцитов:
наименьшее количество отмечалось при стрессе, несколько большее —
при реакции тренировки и наиболее высокое — при реакции спокойной
и повышенной активации (Клетке Г. Э., 1989). Благоприятное влияние
реакций тренировки и особенно активации отмечалось и при исследовании
ряда показателей клеточного и гуморального иммунитета у больных раком
гортани и полости рта (Би-рина Л. М., Гаркави Л. X., Кваки-на Е. Б.,
Кузьменко Т. С, Чиж Г. И., 1987).
В подавляющем большинстве случаев мы отметили четкую корреляцию между
от-носительным числом лимфоцитов в периферической крови и
противоопухолевым эффек-том, т. е. уровнем противоопухолевой
резистентности организма. Вместе с тем в единич-ных случаях, особенно
при слишком высоком лимфоцитозе, такой четкой корреляции не было.
Возможно, это объясняется тем, что РПА переходит в переактивацию
— состояние, далекое от нормы, при котором относительное число
лимфоцитов выше, чем при РПА. Иногда переактивация может
по лейкограмме выглядеть как
активация. Это
62
бывает при наличии нескольких выраженных признаков напряженности в
лейкоцитарной формуле, когда высокий лимфоцитоз как бы маскируется
— при выведении процентного числа лимфоцитов оно оказывается
заниженным и формально соответствующим норме реакции активации. Наличие
лейкопении, характерной для переактивации при опухолевом процессе,
говорит в пользу этого предположения. В подобных случаях особенно важно
определить качественные характеристики лимфоцитов как по широко
известным иммуно-логическим критериям, так и различным биохимическим, в
том числе ферментной актив-ности, меняющейся при разных функциональных
состояниях организма (Нарциссов Р. П., Комиссарова И. А., 1973;
Яновская А. С, 1973; Шепотиновский В. И., Микяшинович 3. И., 1980).
Использование реакции активации совместно с различными препаратами
(циклофосфан, тиоТЭФ, сарколизин, оливомицин) или облучением, а также с
комбинацией этих препара-тов и облучения приводило к защите животных с
опухолями от повреждающего действия таких видов лечения не только без
снижения его противоопухолевого эффекта, но даже с его повышением. В
опытах с введением сарколизина крысам с саркомой-45 было получе-но не
только уменьшение лейкопении, вызываемой сарколизином (в дозе 2 мг/кг
ежеднев-но; суммарная доза 42 мг/кг), но и увеличение
противоопухолевого эффекта: процент торможения в контрольной группе
(только сарколизин) —44, в опытной—76. Исследова-ние
тимико-лимфатической системы показало, что масса тимуса и его
гистологическая картина у крыс с введением сарколизина, несмотря на
торможение роста опухоли, были характерными для стресса (42,0
±11,7 мг), а в лимфоузлах отмечались деструктивные про-цессы, в
то время как у животных с введением сарколизина при поддержании реакции
ак-тивации тимус был увеличен (186±42,3 мг), а в лимфоузлах
наблюдались процессы про-лиферации. При этом возрастало число
бляшкообразующих клеток: 1,34±0,16 по сравне-нию с
0,27±0,02 при стрессе (Р<0,05). Совместное действие
химиопрепаратов и облучения на крыс с опухолями вызывало еще более
глубокий стресс, чем у крыс без опухолей: на-рушение функциональной
активности коры надпочечников, функции щитовидной железы, уменьшение
тимуса, числа лимфоцитов и развитие лейкопении. Применение
активирую-щих воздействий переводило стресс в реакцию активации и,
таким образом, при одновре-менном увеличении противоопухолевого
эффекта, защищало организм от повреждающего действия (Гаркави Л. Х.
Колесникова В. А., Андреева Л. А., 1971; Огородникова Л. С, Гаркави Л.
X., Андреева Л. А., 1973; Уколо ва М. А., Гаркави Л. X., Квакина Е. Б.,
1973; Квакина Е. Б., Гяр-кави Л. X., Уколова М. А., Тарасьянц Ф. М.,
Мулатова А. К., Григорьева Е. Г., 1980; Тарасьянц Ф. М., Гаркави Л. X.,
Квакина Е. Б.,
63
Мулатова А. К., Уколова М. А., 1982; Захарюта Ф. М., 1989).
Антистрессорный характер изменений при развитии реакции активации,
высокий (в пределах верхней половины зоны нормы) уровень
функционирования регуляторных и защитных систем организма, свойственный
обычно молодому возрасту, способность реакции активации влиять на
провоспалительный потенциал организма, ее нормализующее действие на
систему свертывания крови (антикоагуляционное действие), быстрое и
значи-тельное повышение неспецифической и, в том числе,
противоопухолевой резистентности — все это делает реакцию
активации чрезвычайно полезной для практики. Хорошо из-вестно, что
большинство самых различных заболеваний в той или иной мере связано со
снижением резистентности организма. Поэтому целенаправленное повышение
резистент-ности должно быть полезным в комплексе лечебных мероприятий.
Реакция активации на-ходит широкое применение в лечении самых различных
заболеваний. Кроме того, реакция активации может быть использована для
быстрого повышения устойчивости организма к повреждающим факторам и
нагрузкам самой различной природы (например, в спортивной и космической
медицине).
Реакция активации может быть успешно использована с целью профилактики
забо-леваний, так как с ее помощью удается выводить организм из
состояний предболезни, что крайне важно для снижения заболеваемости и
является особенно актуальным. Можно на-деяться, что реакция активации
будет очень полезна для массового оздоровления населе-ния.
О ПРИНЦИПЕ ПЕРИОДИЧНОСТИ В РАЗВИТИИ АДАПТАЦИОННЫХ РЕАКЦИИ И АРЕАКТИВНОСТИ
В предыдущих главах показано, что в основе приспособительной
деятельности ор-ганизма лежит количественно-качественный принцип: на
различное количество раздражи-теля организм отвечает различным
качеством приспособительных реакций. Независимо от качества
раздражителя в ответ на слабые раздражители развивается реакция
тренировки (Гаркави Л. X., 1969; Квакина Е. Б., Уколова М. А., 1969),
на раздражители средней силы — реакция активации (Гаркави Л. X.,
1968, 1969), а на сильные — реакция стресс, опи-санная ранее Г.
Селье.
Периодическая закономерность развития
общих адаптационных реакций
Необходимо отметить, что реакция тренировки, активации и даже стресс
вызыва-лись в наших опытах сравнительно небольшими величинами
раздражителей. Используе
64
мая для электрических раздражений гипоталамуса через хронически
вживленные электро-ды сила тока была сравнительно невелика.
Тренировочной оказалась сила тока, вызываю-щая первую заметную реакцию
организма (чаще всего, ориентировочную). У разных жи-вотных сила тока
колебалась от 0,05 до 0,3 мА. Активирующими оказались раздражения,
вызывающие общую четкую двигательную реакцию животных: сила тока
0,5-1,5 мА. Стрессорным считалось раздражение, вызывающее общие
судороги,—2—5 мА.
Для фармакологических нейротропных веществ и биостимуляторов это были
дозы меньшие, чем обычные терапевтические.
Так, дозы адреналина (подкожные инъекции), необходимые для
получения реакции тренировки у крыс, составляли 0,05 — 0,1 мл
0,001 %-го раствора, для получения реакции
активации—0,05—0,1 мл 0,01%-го раствора и стрессорные
дозы—0,3—0,5 мл 0,01%-го раствора. У человека реакцию
тренировки вызы-вали адреналином в дозах меньших, чем 0,1
терапевтической дозы (0,02—0,1 мл 0,1 %-го
раствора), реакцию активации — дозой в
4—10 раз меньше
терапевтической (0,1-0,25 мл 0,1%-го раствора адреналина).
Из биостимуляторов, например, мумие чаще всего вызывало реакцию
тренировки при скармливании крысам в дозах до 2 мг, реакцию активации
— от 2 до 4 мг, реакцию стресс — более 4 мг. У людей
реакцию тренировки вызывали при однократном приеме 30—70 мг,
реакцию активации —70—120 мг — дозы, значительно
меньшие предлагаемых для терапевтических целей (Шакиров А Ш.. 1976).
Магнитные поля мы применяли в диапазоне магнитной индукции 10—50
мТ, час-тотой 50 Гц в прерывистом (2 с поле, 2 с пауза) и непрерывном
режимах, в течение 5—20 мин (аппарат «Полюс»
ВНИИМПа), а также при эксцентричном вращении постоянных магнитов в
течение 5 — 20 мин. Реакцию тренировки получали, главным образом,
при действии постоянных (неподвижных) магнитов и переменного МП
аппарата «Полюс» в непрерывном режиме с индукцией
10—20 мТ, реакцию активации — при действии пере-менных
полей, чаще всего аппарата «Полюс» в прерывистом режиме с
индукцией 20—40 мТ. Известно, что в биологических исследованиях
использовались МП, по интенсивности во много раз превышающие величины,
применяемые нами. Однако МП такой большой интенсивности не отличались
ни особо высокой биологической активностью, ни выра-женным повреждающим
действием.
Анализируя данные литературы об изменениях в организме при действии
самых различных биологически активных факторов (в широком диапазоне
доз), можно заклю-чить, что эти изменения укладываются в картину
описанных адаптационных реакций. Од-нако во всем диапазоне действующих
доз — от минимально действующей
65
до максимальной, смертельной — такой простой
количественно-качественной зависимо-сти: слабые воздействия —
реакция тренировки, большие по величине — реакция актива-ции,
сильные, чрезвычайные—реакция стресс — отметить не удается.
Напротив, нередки случаи, когда изменения, характерные для реакции
тренировки, возникают при действии довольно больших величин (доз)
раздражителей, в то время как стресс — при действии сравнительно
небольших величин (доз). Изменения, характерные для реакции активации,
возникают и при действии сравнительно небольших доз, и довольно больших
доз, и про-межуточных между ними. Встречается и такой вариант: какая-то
доза раздражителя вызы-вает развитие изменений, характерных для
стресса, а увеличение ее приводит не к углуб-лению стресса, истощению и
гибели организма (согласно Г. Селье), а как бы к смягчению изменений
вплоть до полного их отсутствия.
Как объяснить эти факты с точки зрения наших представлений? Конечно,
для суж-дения о силе воздействия на живой организм недостаточно знать
абсолютную величину (силу, дозу) действующего фактора. Биологическая
активность определяется не только этой абсолютной величиной, но и
чувствительностью организма. Может быть, все дело в различной
чувствительности?
Однако, как ни заманчиво такое объяснение, его нельзя признать
единственно пра-вильным. Разница в индивидуальной чувствительности,
конечно, может быть значитель-ной, но все же не может дать ключ к
пониманию того, почему стресс — реакция на силь-ный раздражитель
— у животных одного и того же вида может вызываться величинами на
порядки меньшими, чем реакция тренировки — реакция на слабый
раздражитель. Кроме того, в процессе длительного поддержания в
организме реакции тренировки или актива-ции дозу действующего фактора
приходилось соответственно либо волнообразно увели-чивать, либо также
волнообразно уменьшать. Если же несколько дней применялась одна и та же
доза, то происходило чередование реакций, а для поддержания одной
реакции дозу (силу) приходилось корригировать. Для коррекции дозу
необходимо было изменять всего лишь на 10—20%, чаще на 20%. Иными
словами, в эксперименте для перехода из одной реакции в другую
(соседнюю) необходимо изменять количество действующего фактора всего
лишь на 20%. Таким образом, основные реакции — тренировки,
спокойной и повы-шенной активации и стресса — можно было вызвать
в узком диапазоне дозы (силы) дей-ствующего фактора. В связи с этим у
нас возникло предположение, что в организме суще-ствуют различные
уровни реактивности, диапазоны (этажи). На каждом из этих уровней
существуют четкие количественно-качественные отношения: слабые для
данного уровня раздражители вызывают реакцию тренировки, средние
— реакцию спокойной и повы-шенной активации, сильные —
стресс. Эти тетрады адаптационных реакций повторяются
66
на каждом уровне. Реактивность — это, фактически, способность
отвечать реакцией. Если организм отвечает реакцией на действующий
фактор, малый по абсолютной величине, это свидетельствует о высоком
уровне реактивности (низкий этаж). Фактически, живой орга-низм может
«работать» на разных уровнях реактивности, Если однотипная
реакция спо-собна развиваться в ответ на действие разных по абсолютной
величине раздражителей, это значит, что она развивается на разных
уровнях реактивности (этажах).
Для проверки этого предположения мы исследовали влияние на организм
различ-ных по качеству раздражителей (адреналин, магнитное поле, мумие)
в широком диапазоне доз, последовательно уменьшая или увеличивая дозы
по сравнению с применяемыми ра-нее (на 10—20%). При этом
изучались некоторые показатели, характеризующие адапта-ционные реакции.
Исследования показали, что по мере увеличения (или уменьшения) дозы
раздражи-телей действительно происходит повторение основных
адаптационных реакций, триад, как мы считали ранее,— тренировки,
активации, стресса (Гаркави Л. X., Квакина Е. Б., 1975—1986;
Квакина Е. Б., Гаркави Л. X., 1975), и тетрад (рис. 37) —
тренировки, спокой-ной активации, повышенной активации, стресса
(Гаркави Л. X., Квакина Е. Б., 1987). К настоящему времени нами
обнаружено более десяти уровней реактивности (диапазонов, этажей). Пока
трудно сказать, сколько этажей может существовать на уровне организма,
так как если нижняя граница реактивности (высокий этаж) ограничивается
летальной до-зой (силой) действующего фактора, то границу вниз по дозе
(силе) нам пока определить не удалось. Возможно, для многих факторов
(например, кислород, углекислота, магнитное поле) эта граница также
будет связана с летальным эффектом, обусловленным не избыт-ком, а
недостатком фактора, так как организм реагирует на изменение в любую
сторону. Применение раздражителей, превышающих максимальную
(стрессорную) для данного уровня (диапазона, этажа) интенсивность,
приводит к выключению этого диапазона и включению следующего, в
пределах которого раздражители также различаются в соответствии с их
силой, однако параметры их уже совершенно иные: слабый раздражитель для
последующего диапазона (этажа) превышает величину сильного раздражителя
предыду-щего. Сила раздражителя является относительной величиной:
реакция развивается как бы не на раздражитель целиком, а на приращение
величины этого раздражителя (по сравне-нию с нижележащим уровнем
реактивности).
Таким образом, организм имеет как минимум две шкалы отсчета силы
воздействия: одну — относительную (относительно данного уровня
реактивности), определяющую ха-рактер развивающейся адаптационной
реакции, другую — абсолютную, определяющую уровень реактивности.
67
Рис. 37. Периодическая смена тетрад адаптационных реакций: тренировки
(1), спокойной активации (2), по-вышенной активации (3), стресса (4)
Нелинейность изменений (по многим показателям реакций), отражающая
дискрет-ность реакций в пределах одного уровня реактивности (рис. 38),
закономерно повторяется на каждом уровне реактивности и особенно при
переходе с одного уровня на другой.
Периодичность в развитии адаптационных реакций, по нашему мнению,
позволяет организму, с одной стороны, гибко приспосабливаться даже к
небольшим изменениям факторов среды (уже на 10—20%), а с другой
— расширяет границы воздействий, в усло-виях которых организм
остается жизнеспособным. Иными словами, периодически повто-ряющиеся
тетрады адаптационных реакций составляют реальный механизм системы
мно-гоуровневой регуляции гомеостаза. Можно только удивляться, какая
гибкая количествен-но-качественная система адаптации создалась в
процессе эволюции живого. Изучение ко-личественных соотношений силы
(дозы) воздействия, вызывающих развитие адаптацион-ных реакций на
разных уровнях, показало, что независимо от уровня (этажа) для полу-
68
Рис. 38. Нелинейные изменения некоторых показателей при увеличении силы
раздражителя, вызывающего последовательно реакции тренировки, активации
и стресса:
а — нелинейные изменения глюкокортикоидов (1) и массы тимуса (2);
б— нелинейные изменения электрического порога раздражения нервных
структур гипоталамуса при граду-альном увеличении относительного числа
лимфоцитов от реакции стресс к реакции активации.
чения каждой следующей реакции предыдущую дозу нужно увеличить, умножив
на один и тот же коэффициент (К)—шаг между реакциями. У здоровых
людей и животных К ча-ще всего равен 1,2— 1,25, но может
доходить до 1,3. У больных, особенно тяжелоболь-ных.
69
К уменьшается вплоть до 1,1. При старении К также уменьшается.
Таким образом, была получена
логарифмическая зависимость между величиной воздействия и
ответной адаптационной реакцией в большом диапазоне силы (дозы).
По-явление одной и той же реакции происходило при уменьшении
(увеличении) дозы в К4 раз. Логарифмическая зависимость между величиной
раздражения и реакцией хорошо из-вестна в физиологии. Она лежит в
основе так называемого психофизического
закона (закона Вебера — Фехнера), описывающего
соотношение между величиной раздражения и ощущением. Изменение
ощущения происходит также при изменении силы раздражения на
10—20%. Иными словами, организм логарифмирует раздражители на
разных иерархи-ческих уровнях.
Принцип периодичности в развитии адаптационных реакций позволяет
объяснить механизм действия на организм раздражителей очень малой
величины (вплоть до гомео-патической), нелинейный
характер зависимости
эффекта от
интенсивности воз-действия, а также большую эффективность
меньших по абсолютной величине воздейст-вий. Особенно убедительно эти
положения иллюстрируются работами из области магни-тобиологии (Плеханов
Г. Ф., 1967; Прес-ман А. С, 1968; Музалевская Н. И., 1973; Муза-левская
Н. И., Шушков М. Д., 1978; Макеев В. Б., 1979; Акоев И. Г., 1983, 1986;
Жуков Б. Н., Труфанов Л. А.,
Мусиенко Е. Н., 1980; Красногорская
Н. В., Сперанский А. П., Десницкая М. М. 1984; Макеев В. Б., Темурьянц
Н. А., Владимирский Б. М., Тишкина О. Г., 1984), возможно потому, что
МП применяются и в эксперименте, и для лечения в
зна-чительно больших диапазонах, чем,
например, другие, физиотерапевтические или, тем более,
фармакологические средства. Нами также получены различные адаптационные
ре-акции при использовании переменных магнитных полей с
индукцией около 0,1 мТ (Гар-кави Л. X., Квакина Е. Б., Уколова М.
А., 1978; Гаркави Л. X., Квакина Е. Б., Коробейни-кова Е. П., Уколова
М. А., Шихлярова А. И. и др.,
1984).
Однако такая же закономерность сохраняется и для лекарствен-ных
средств. О биологической активности очень малых доз (на примере брома и
кофеина) четко говорил И. П. Павлов: «Не подлежит сомнению, что
дозировка имеет гораздо большее значение вниз, чем вверх. Вся штука в
варьировании дозировки вниз... Вы должны обратить внима-ние на строгую
дозировку — уменьшать дозу, а не увеличивать. Все наши
«собачьи» опы-ты говорят за это» (Павловские
клинические среды. М., 1954. Т. 1. С. 79).
Известны работы А. Н. Кудрина, свидетельствующие о фармакологическом
влия-нии чрезвычайно малых доз биологически активных веществ, например
адреналина в дозе 1.10-22 г/мл (Батрак Г. Е. Кудрин А. Н., 1979; Кудрин
А. Н., 1977), причем интересно, что отмечает несколько оптимумов
на уровне разных концентраций
70
адреналина («ступенеобразная
закономерность», по определению
авторов).
Болгарский фармаколог В. Петков (1974) указывает на активность
адреналина в до-зах в 1000—10000 раз меньших, чем терапев
тические, причем в этих случах адреналин может вызывать даже снижение
кровяного давления и гипогликемию. Наши исследования показали, что
адреналин в таких дозах вызывает развитие гармоничной реакции
актива-ции как спокойной, так и повышенной. У людей подобные дозы
одновременно с развити-ем реакции активации вызывают снижение
артериального давления и содержания сахара в крови до уровня нижней
половины зоны нормы.; Развитие общей реакции в ответ на вве-дение таких
малых доз можно объяснить, во-первых, тем, что эти дозы находятся в
зоне физиологических значений, и, во-вторык, тем, что организм способен
реагировать, как уже говорилось, на изменение не только вверх, но и
вниз. Отсюда понятно положение М. Н. Кондрашовой «о сильном
влиянии слабых воздействий». Фактически речь идет о сиг-нальных
информационных воздействиях, которые хотя и являются значительно более
сла-быми по сравнению с энергетическими, но обладают не меньшим, а в
ряде случаев — большим действием на организм животных и человека.
Это объясняется тем, что по мере усложнения природных объектов роль
информационных взаимодействий (родовых фак-торов) увеличивается
(Николаев Л. А., 1976), более того — они становятся управляющими
(Николаев Л. А., 1984). Иными словами, высокоорганизованным системам
присущи энер-гетически неэквивалентные взаимодействия (Сетров М. И.,
1975).
А. С. Пресманом (1968) была выдвинута концепция об информационной
функции электромагнитных полей (ЭМП) в жизнедеятельности
организмов, причем имелась в ви-ду передача информации из внешней среды
в организм, информационная взаимосвязь внутри организма и обмен
информацией между организмами. Эта концепция основыва-лась на фактах,
которыми располагала к тому времени магнитобиология: 1) наличие
раз-личного рода влияния на организм естественных и очень слабых
искусственных ЭМП, когда сколько-нибудь заметные энергетические
эффекты полностью исключаются; 2) за-висимость характера
реакции организма не от величины электромагнитной энергии, по-глощаемой
в тканях, а от модуляционно-временных параметров ЭМП, причем величина
реакции зачастую по мере увеличения интенсивности ЭМП даже
уменьшалась. Возни-кает вопрос, в чем
сходство и в чем различие одной и той же адаптационной реакции,
раз-вивающейся на разных уровнях реактивности? Процентное число
лимфоцитов остается характерным для типа реакции на разных уровнях
реактивности. Однако другие показатели лейкоцитарной формулы в
значительной степени помогают ответить на вопрос, происходит ли
развитие реакции организма на
71
низких или высоких уровнях. Так, при реакциях высоких уровней
реактивности (низкие этажи, малые по абсолютной величине воздействия)
деятельность подсистем организма почти всегда бывает гармоничной,
согласованной, и все элементы лейкоцитарной форму-лы находятся в
пределах нормы. Когда же нагрузка по абсолютной величине увеличивает-ся
(низкие уровни реактивности, высокие этажи), гармоничность
работы подсистем ор-ганизма
часто нарушается, что находит отражение в
лейкоцитарной формуле,— появ-ляются отклонения от нормы в
содержании ее элементов либо в сторону повышения, либо в сторону
снижения. Отмечаются моноцитоз или монопения, эозинофилия или
эозинопе-ния, вплоть до полного отсутствия эозинофилов, сдвиг
влево (увеличение числа палоч-коядерных нейтрофилов), увеличение
числа базофилов. Кроме того, происходит либо
уменьшение (лейкопения), либо
увеличение общего числа лейкоцитов (лейкоцитоз). Эти
отклонения мы называем «элементами напряженности»
реакций. Чем больше эле-ментов напряженности, тем больше оснований
считать, что это реакции более высокого этажа (низкого уровня
реактивности). Появление элементов
напряженности говорит о рассогласовании,
десинхронизации деятельности какой-либо
из подсистем организма. Так,
например, отсутствие эозинофилов (анэозинофилия)
свидетельствует об избыточ-ности секреции глюкокортикоидов для данной
адаптационной реакции; напротив, избы-ток
эозинофилов (аэозинофилия) — о
недостаточности секреции глюкокортикоидов для данной реакции.
Эозинофилия на фоне реакции активации
(процентное, содержание лимфоцитов в пределах верхней половины зоны
нормы) свидетельствует об относительной недостаточности секреции
глюкокортикоидов, а эозинофилия на фоне стресса (процентное
содержание лимфоцитов ниже нормы) — об абсолютной недостаточности
глюкокортикоидной секреции, т. е. о наступившей стадии истощения
стресса (Гаркави Л. X., Мацанов А. К., 1973).
Наличие не одного, а нескольких признаков напряженности говорит о более
резко выраженной рассогласованности работы подсистем организма и
встречается при адапта-ционных реакциях на довольно высоких этажах.
Если признаки напряженности являются стойкими, повторяются при анализах
в течение длительного времени и мало поддаются коррекции при
воздействиях, это свидетельствует о стойком нарушении работы в
какой-либо из подсистем организма. В этих случаях даже реакции
повышенной и спокойной ак-тивации и тренировки нельзя назвать
физиологическими.
Признак напряженности по основному показателю реакций —
процентному содер-жанию лимфоцитов — говорит при снижении (ниже
уровня нормы) о развитии стресса, а при избыточном повышении — о
развитии состояния так называемой переактивации. И стресс, и
переактивация являются неспецифической основой раз-
72
вития многих заболеваний: стресс — одних, а переактивация —
других. Стойкий стресс со многими элементами напряженности и
переактивация обычно свидетельствуют о том, что реакции развиваются на
низких уровнях реактивности (высоких этажах).
Соотношение глюкокортикоидных и минералокортикоидных гормонов коры
над-почечников и функциональной активности щитовидной железы также
осталось характер-ным для каждой из адаптационных реакций: при стрессе
отмечается резкое преобладание глюкокортикоидов
при низком уровне
секреции минералокортикоидов и снижении функциональной
активности щитовидной железы; при реакции активации —
высокий уровень минералокортикоидов при нормальном
или слегка повышенном (реакция повы-шенной активации) уровне
глюкокортикоидов и повышенной до верхних границ нормы функции
щитовидной железы; при реакции тренировки — умеренное повышение
глюко- и минералокортикоидов и функциональной активности щитовидной
железы. Тимус умень-шен при развитии стресса, увеличен при реакции
активации и занимает промежуточное положение при реакции тренировки.
Однако при реакции активации как спокойной, так и повышенной,
развивающейся при действии малых доз, тимус увеличен больше. Так,
на-пример, при реакции спокойной активации на высоких уровнях
реактивности (низких этажах) масса тимуса крыс составляла
392,4±10,1 мг, на низких — 207,8±9,2 мг, при
реак-ции повышенной активации тимус на высоких уровнях реактивности
доходил до 500 мг. При стрессе высоких этажей угнетена вся
тимико-лимфатическая система. По мере уменьшения абсолютной
величины раздражителя, вызывающего стресс других уровней, уменьшается
угнетение лимфоидной ткани тимуса. Например, при стрессе низких
уровней (высокие этажи) масса тимуса составляла
38,6±7,8 мг, при развитии стресса на высоких
уровнях реактивности — 71,2 ± 9,1
мг. Затем уменьшается
угнетение лимфоидной ткани
селезенки; выраженное угнетение остается лишь
на уровне лимфоузлов. Кроме того, отмечается перераспределение
активности гормонов. При развитии всех реакций вы-соких этажей
увеличивается секреция, глюкокортикоидных гормонов и снижается —
ти-реоидных и половых. На низких этажах наблюдаются обратные отношения.
Эти измене-ния происходят в рамках колебаний величины секреции. Так,
например, при тяжелом стрессе, вызванном большим по абсолютной величине
воздействием (низкие уровни реак-тивности), содержание кортизола
доходит до 612±70,5 нмоль/л, в то время, как
при бо-лее мягком стрессе (высокие уровни реактивности) —
406±51,3 нмоль/л.
В литературе, посвященной
исследованиям по стрессу, часто
встречаются термины: «умеренный» стресс,
«мягкий» стресс и т. п., т. е. ощущается потребность
ко-личественной оценки раздражителей, вызывающих стресс. Такие
термины иногда под-крепляются конкрет-
73
ными данными. Так, по мере увеличения интенсивности стресса описано
вовлечение в ре-акцию сначала системы блуждающий нерв— инсулин,
затем — симпатико-адреналовой системы и системы гипофиз —
кора надпочечников и, наконец, системы блуждающий нерв— глюкагон
(Сох Т., 1978). С другой стороны, при стрессе иногда описывают
изме-нения в организме, которые, по нашим представлениям, могут
наблюдаться при высоко-этажном стрессе, например, когда говорится не
только о снижении секреции тестостерона, но и о полной его блокаде
(Mason J, 1975).
С целью поисков показателя уровня (этажа), на котором развивается
адаптационная реакция, а также поисков простого,
«бескровного» показателя реакции, проводилось ис-следование
электрического потенциала кожи (ЭПК) и аутофлоры.
У обследованных лиц (130 человек) по анализу лейкограммы устанавливался
тип адаптационной реакции (активация, тренировка или стресс) и
параллельно проводилось измерение потенциала в избранных акупунктурных
точках (12 точек-источников и 8 точек переднего и заднего срединных
меридианов).
Исследование показало, что значения потенциалов в изучаемых точках
охватывали диапазон от —80 мВ до +200 мВ. Наибольшая взаимосвязь
между значением потенциала и типом адаптационной реакции по показателям
крови была установлена в точке J. Харак-тер взаимосвязи проявлялся в
виде периодического повторения по шкале значений потен-циала диапазонов
по 15 мВ, причем на долю реакций тренировки и активации приходи-лось
соответственно по 5—6 мВ, а стрессу соответствовал внутри каждого
диапазона ин-тервал в 3 мВ. Это и послужило основой для определения
типа адаптационной реакции организма по измерению потенциала в точке J.
Между рассматриваемыми классификаци-онными признаками —
лейкограммой и потенциалом точки J—была обнаружена сильно
выраженная, статистически надежная взаимосвязь (по критерию х2 и
коэффициенту со-пряженности Пирсона). Была проанализирована возможность
использования метода из-мерения потенциала точки I в качестве теста для
массовых обследований населения. Ока-залось, что предложенный тест
определения общей неспецифической адаптационной ре-акции по потенциалу
данной акупунктурной точки обеспечивает высокую статистическую
надежность распознавания реакции активации (коэффициент прогнозирования
положи-тельного результата 0,863) и реакции тренировки (0,867), но
низкую надежность распо-знавания реакции стресс (0,412), т. е. для
выявления стресса необходимы дополнительные критерии оценки
(Самохвалова Л. И., Дерибас В. И., Белокопытов В. И., 1987).
Аутофлора кожи является одним из показателей неспецифической
резистентностй организма, поэтому мы предположили, что этот показатель
может быть информативным для суждения о ха-
74
рактере развивающейся адаптационной реакции и, возможно, для суждения
об уровне ре-активности (этаже),[на котором эта реакция развивается.
Было найдено, что при стрессе аутофлора кожи самая обильная, особенно
велико число «желтых» маннитпозитивных колоний —
патогенных форм. При реакции тренировки число «желтых»
колоний меньше, чем при стрессе. Наименьшее число колоний, вплоть до
полного их отсутствия, отмечает-ся при реакции активации.
Характер и количество микрофлоры кожи зависели не только от типа
адаптацион-ной реакции, но и от уровня реактивности, на котором она
развивалась. Необходимо еще раз подчеркнуть, что реакции тренировки,
активации и стресса развиваются не только при нашем вмешательстве, но
существуют независимо от него, как и уровни реактивности. Так, у
тяжелых или ослабленных длительной болезнью людей развиваются стресс
или ре-акции тренировки и активации низких уровней реактивности
(Гаркави Л. Х., Квакина Е. В., 1987). Исследование аутофлоры при
стрессе низких уровней (высоких этажей), напри-мер, у больных с
хроническими воспалительными процессами, выявило обильную пато-генную
микрофлору (49,5± ±4,2 желтых колоний), в то время как
при стрессе высоких уровней реактивности (состояние
«предболезни» ) количество патогенной микрофлоры было
существенно меньше (8,0 ± 1,7, р<0,01). Кроме того, при
стрессе низких уровней были обнаружены гемолитические формы
стафилококков и стрептококков. У больных и ослабленных людей при
наличии реакций тренировки и активации с признаками напря-женности по
лейкограмме (низкие уровни реактивности), количество желтых колоний
бы-ло существенно больше, чем при одноименных реакциях высоких уровней
реактивности: при реакции тренировки 26,0 ± 2,5 и 5,5 ±
2,8 соответственно, р<0,01; при реакции актива-ции
—4,5±1,5 на низких уровнях при почти полном отсутствии
— единичные колонии — на высоких. Эти данные говорят в
пользу того, что на нижних диапазонах (этажах) неспе-цифическая
резистентность организма повышена больше. Для использования ЭПК и
ау-тофлоры кожи в качестве показателей адаптационных реакций нужны
дальнейшие иссле-дования. Разумеется, необходимы также поиски других
простых и доступных, показателей адаптационных реакций и уровней, на
которых они развиваются.
Как уже неоднократно говорилось, простым и доступным показателем
адаптацион-ной реакции, используемым при подборе дозы (силы)
воздействия, является лейкоцитар-ная формула. Однако, так как в
зависимости от уровня реактивности процентное содержа-ние лимфоцитов,
характеризующее каждую реакцию, не меняется, этот показатель может быть
использован для суждения о том уровне, на котором развивается данная
реакция, только в том случае, если изменение реакции соответствует
изменению применяемой до-зы (силы). Когда развитие адаптационной
реакции обусловлено дейст-
75
вием специального раздражителя, мы судим об уровне (этаже)
по величине (дозе, силе) использованного
раздражителя, в том случае, если изменения реакции
соответствуют изменению дозы
(силы). Иными словами, проводится своеобразная
функциональная проба работы определенного уровня
реактивности. Если же применяемое воздействие не
вызывает развития ожидаемых реакций, это означает, что данный уровень
реактивно-сти не работает. Чаще всего при этом дозу (силу) приходится
увеличивать. Когда же реак-ция развивается без специального
воздействия, об уровне реактивности мы судим по от-сутствию или наличию
и степени выраженности элементов напряженности. У людей, кроме того, об
уровне реактивности можно судить по самочувствию. Даже реакции
тре-нировки и активации на высоких этажах могут сопровождаться
какими-либо нарушения-ми самочувствия. В результате исследований было
показано, что при заболеваниях реак-ции всегда развиваются на низких
уровнях реактивности, причем, чем тяжелее заболева-ния, тем ниже
уровень реактивности (выше этаж). По мере старения организм также
переходит на более низкие уровни реактивности.
Большой интерес представляет изучение энергетического обмена, так как
знание энергетической цены резистентности при развитии адаптационных
реакций на разных уровнях позволяет избрать в каждом случае оптимальный
уровень, а также необходимое биохимическое сопровождение для
оптимизации энергетического обмена (Кондрашова М. Н., 1975).
Исследования, проведенные совместно с М. Н. Кондрашовой, Е. И.
Маевским, Е. А. Косенко и Ю. Г. Каминским, показали, что
энергетические траты на уровне малых доз значительно меньше, чем на
уровне больших доз.
Энергетический обмен при одноименных реакциях разных уровней
реактивности сохраняет, в той или иной степени, присущие данной реакции
черты. Так, при реакции ак-тивации фонд адениловых
нуклеотидов (АДФ + АТФ)
и концентрация фосфоенол-пирувата (ФЕП),
отражающего вклад энергетического обмена в биосинтез
высокомолеку-лярных соединений в ткани печени, наиболее высоки. Реакции
дыхательной цепи мито-хондрий свидетельствуют об интенсификации
энергодающего окисления и воспроизвод-ства эндогенного
сукцината (Косенко Е. А., Каминский Ю. Г., Кондрашова
М. Н., Маев-ский Е. И., Окон Е. В., Тарасьянц Ф. М., Котляревская Е. С,
Барсукова Л. П., 1978). При увеличении абсолютной величины дей- ;
ствующего фактора, т. е. на низких уровнях ре-активности, появляется
регуляторное ограничение гиперактивного
окисления, повиди-мому, обусловленное щавелево-уксусной
кислотой (Григоренко Е. В., Кондрашова
М. Н., 1986; Барсукова Л. П.,
Котляревская Е. С, Марьяновская Г. Я., 1987). Иногда наблю-даются
признаки нерезко выраженного низкоэнергетического сдвига. Следовательно,
76
реакция активации, названная М. Н. Кондрашовой (1979), «золотой
серединой» (по энер-гетическим характеристикам), является такой
лишь на высоких уровнях реактивности; при влиянии малых по абсолютной
величине действующих факторов.
Состояние митохондрий, характерное для реакции активации, по
классификации М. Н. Кондрашовой и др. (1987), можно обозначить как
гиперактивное. При переактива-ции, которая встречается на низких
уровнях реактивности (высоких этажах) в отличие от физиологической
активации, скорость дыхания «после работы» существенно
превышает начальную скорость дыхания — признак
низкоэнергетического
сдвига.
При реакции тренировки, которая отличается наибольшим накоплением фонда
ян-тарной кислоты (ЯК), в ответ на повышение величины действующего
фактора (увеличе-ние этажа, снижение уровня реактивности) отмечается
лишь увеличение скорости фосфо-рилирующего дыхания. Щавелево-уксусное
ограничение гиперактивной СДГ и признаки низкоэнергетического сдвига
почти отсутствуют. По-видимому, преобладание накопи-тельных процессов
при этой реакции позволяет на какое-то время на высоких этажах не иметь
признаков гиперактивного напряженного состояния.
При стрессе со снижением уровня реактивности появляется более глубокое
тормо-жение — патологическое ингибирование гиперактивной СДГ
продуктами окисления ад-реналина и
серотонина (Григоренко Е. В., Кондрашова М. Н., 1987). Фонд
эндогенной ЯК и его воспроизводство незначительны. При тяжелом стрессе
низких уровней реактив-ности может отмечаться существенное снижение
скорости окисления ЯК, резкое увеличе-ние времени фосфорилирования при
достаточно высоком фонде эндогенной ЯК. В этом случае возможно
«аварийное» переключение энергетической регуляции с
углеводного на липидный источник энергии . (Гаркави
Л. X., Квакина Е. Б., Барсукова Л. П., Котлярев-ская В. С,
Марьяновская Г. Я., 1983, 1984; Барсукова Л. П., Котляревская Е.
С, Марьянов-ская Г. Я., 1987). При стрессе, особенно
с увеличением этажа, повышается начальная
скорость дыхания. При этом скорость дыхания
«после работы» также растет и превыша-ет начальную
скорость. Состояние энергетики при стрессе укладывается в
картину «ги-перактивного напряженного
состояния» (Кондрашова М. Н. и др., 1987). Необходимо
отметить, что митохондрии стрессорных животных, особенно на низких
уровнях реактив-ности, могут поддерживать интенсивное дыхание
менее продолжительное время, чем ми-тохондрии животных, находящихся в
реакциях активации и тренировки (Барсукова Л. П..
Котляревская Е. С, Марьяновская Г.
Я., 1987). Таким образом, энергетика организма
более экономична на высоких уровнях реактивности, чем на низких.
Изучение энергетики при разных реакциях на разных уровнях
реактивности подтвердило особую роль янтар-ной
77
кислоты, значение которой показано многолетними исследованиями М. Н.
Кондрашовой, в энергообеспечении адаптационной деятельности
организма.
Интересно отметить, что содержание и синтез ФЕП — ключевого
соединения в ре-акциях глюконеогенеза — больше зависит от
характера реакции, чем от уровня реактив-ности, повышаясь максимально
на каждом уровне реактивности при развитии реакции ак-тивации. При этом
увеличение содержания ФЕП отмечается даже ранее, чем увеличение тимуса
и процентного содержания лимфоцитов периферической крови. Концентрация
же глюкозо-6-фосфата больше связана с абсолютной величиной
раздражителя, т. е. уровнем реактивности, и почти градуально отражает
дозу воздействия (Маевский Е. И., Кондра-шова М. H.
Каминский Ю. Г., Косенко Е. А., Гаркави Л. X., Квакина Е. Б., Шихлярова
А. И., 1977).
Как известно, стресс — это реакция, в которой процессы
катаболизма преобладают, что наиболее выражено при стрессе низких
уровней реактивности (высоких
этажей).
Реакция тренировки характерна тем, что в ней мало выражен как распад,
так и син-тез белковых веществ и нуклеиновых кислот. Пока не
удалось установить чёткую зави-симость этих процессов от уровня
реактивности.
При реакции активации высоких и умеренных уровней реактивности
(малые и умеренные дозы) отмечается четкое преобладание
анаболизма — увеличение синтеза белковых веществ и нуклеиновых
кислот, особенно при повышенной активации. Вызов и стойкое поддержание
в течение месяца повышенной активации у
растущих животных (цыплят И крысят) с помощью
влияния ПеМП (0,2 -3,2 мТ, 50 Гц), омагниченной воды и
элеутерококка (жидкого экстракта, разведенного в 10 раз, и добавляемого
дозированно в питьевую воду) приводило к ускорению роста и
увеличению массы животных на 25—30% по сравнению с контролем, т.
е. эти животные становились как бы «великанчиками» —
по терминологии И. А. Аршавского. При этом отмечались изменения,
типичные для повышенной активации (например,
увеличение тимуса и
лимфоидной ткани других органов). Мы думаем, что повышенная
активация умеренных уровней реактивности
со-ответствует состоянию физиологического стресса,
найденного Аршавским (1968—1986). Это принципиально
отличное от стресса Г. Селье одно из физиологических состоя-ний
Аршавскому удалось получить с помощью специально выбранных факторов,
которые в процессе эволюции сформировались как активирующие: мышечное
движение, гипоксия, холод. Их эффективность значительно увеличивается
тем, что они применяются Аршав-ским на начальных стадиях онтогенеза.
При этом используется не чрезмерная, а умерен-ная величина этих
факторов. Поскольку изменения в организме как при повышенной
ак-тивации, так и при «физиологи-
78
ческом стрессе» принципиально отличны и даже во многом
противоположны изменениям, описанным для стресса Г. Селье, нам кажется,
что название «физиологический стресс» — напряжение
— приводит к совмещению несовместимых понятий. С одной стороны,
стресс (напряжение) — неспецифическая реакция на сильный
раздражитель с элементами по-вреждения, десинхронизацией работы
подсистем, преобладанием катаболизма, значитель-ной энтропии и т. п., а
с другой,— прекрасная физиологическая активация организма при
хорошей синхронизации деятельности его подсистем, с преобладанием
процессов анабо-лизма и выраженной негэнтропии. Более четкий
количественный подход помог бы, по нашему мнению, избежать совмещения
разных понятий и подчеркнуть новизну и самостоятельную ценность
открытого Аршавским физиологического
состояния.
В зависимости от уровня реактивности (этажа) происходят и некоторые
изменения активности дегидрогеназ лимфоцитов. При увеличении абсолютной
величины действую-щего фактора (снижение уровня реактивности, повышение
этажа) активность дегидроге-наз лимфоцитов меняется фазно: вначале
увеличивается, а затем уменьшается. Особенно резкое увеличение, а
затем падение отмечается при стрессе, а наименьшее — при реакции
активации. Кроме того, на этапе чрезмерного повышения активности
ферментов увеличивается число жестких корреляционных связей между ними.
Это свидетельствует об увели-чении напряжения энергетических процессов.
На этапе снижения активности, напротив, отмечается резкое ослабление
координации в действии ферментов, что может свидетельствовать об
истощении энергетических ресурсов и снижении адаптационной возмож-ности
клеток.
Интересно также было выявить разницу или сходство в степени повышения
рези-стентности на разных уровнях реактивности. С этой целью была
исследована возможность повышения неспецифической резистентности под
влиянием реакции активации на разных уровнях. Опыты по изучению
неспецифической противоопухолевой резистентности пока-зали, что реакция
активации, вызванная в десятки раз меньшими по абсолютной величине
действующими факторами (адреналин, магнитное поле, мумие), оказывала
даже больший противоопухолевый эффект, хотя эта зависимость нелинейна.
Так, ежедневное введение дозы от 5 до 10 мкг адреналина (на 100 г веса
животного) дало 27% полной регрессии сар-комы-45 и 45% торможения.
Применение 0,6—0,9 мкг адреналина вызвало регрессию в 45,4%
случаев, а торможение — в 18%; 0,12—0,18 мкг —22,2%
регрессии и 44% тормо-жения, а от 0,002 до 0,006 мкг —55%
регрессии и 11% торможения (рис. 39). При воздей-ствии адреналином на
саркому, индуцированную 3,4-бензпиреном, доза 0,6—0,9 мкг
вы-зывала регрессию в 45,5% случаев, а торможение роста — в 27%
случаев;
79
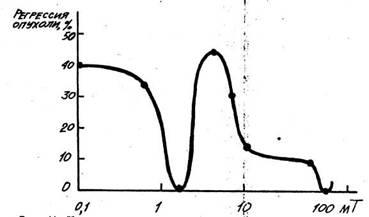
Рис. 39. Характер зависимости величины
противоопухолевого эффекта от дозы (силы)
действующего фактора в широком диапазоне значений (на примере
адреналина)
доза 0,12—0,18 мкг — регрессию в 36,3%,
торможение в 63,3%. При воздействии адре-налином на опухоль
яичника (модель Бискинд) доза 0,12—0,18 мкг приводила в 25%
слу-чаев к регрессии, а в 35% — к торможению роста опухоли; доза
от 0,002 до 0,006 мкг — в 35% случаев — к регрессии
опухолей и в 15%— к торможению роста. При использовании
переменного магнитного поля (аппарат «Полюс-1»,
частота 50 Гц, форма тока синусои-дальная, режим прерывистый— 2 с
поле, 2 с — пауза, ежедневно на голову в течение 2 мин) на модели
саркомы-45 с индукцией 50 мТ полной регрессии опухолей не происходило,
но во всех случаях отмечалось торможение роста. При индукции 35 мТ
регрес-сия опухолей наблюдалась в 10%, торможение — в
40%. 10 мТ —
регрессия 13%, торможение
—38,4%; 3,2 мТ — регрессия 42%, торможение — 58%, при
1,4 мТ полной регрессии опухоли не было, торможение роста наблюдалось в
45% случаев; 0,7 мТ — регрессия 35%,
торможение—50%; 0,1 мТ — регрессия 40%,
торможение — 52% (рис. 40). На модели саркомы,
индуцированной 3,4-бензпиреном,
применялось ПеМП ин-дукции 0,7 мТ. Было
получено: в 13%—полная регрессия опухоли и в 66,6% -торможение
роста. При опухолях яичника ПеМП 0,7 мТ вызывало регрессию в 20%,
торможение—в 60%; 0,1 мТ— регрессию в 30% случаев,
торможение—в 20%. Применение малых доз му-мие (десятые доли
мг) приводило к рассасыванию крысиных сарком в 60—80%
случаев. У остальных животных в одной половине случаев наблюдалось
торможение роста опухо-лей, а в другой —
рост как в контроле. При использовании
в 10 раз большей дозы мумие рассасывание
80
Рис. 40. Характер зависимости величины противоопухолевого эффекта от
дозы (силы) действующего факто-ра в широком диапазоне значений
(на примере магнитных
полей)
наблюдалось в 50% случаев, в 30—-40%'— торможение роста и в 10%—рост (опыты А, И. Шихляровой).
Приведенные результаты опытов говорит о сложном, нелинейном, близком к
вол-нообразному характере зависимости между дозой (силой воздействия) и
противоопухоле-вым аффектом. Во всех сериях опытов при действии и
адреналина, и ПеМП, и мумие пол-ная регрессия опухолей наблюдалась при
вызове и длительном поддержании реакции ак-тивации, торможение
роста — при преобладании в течение времени воздействия реакции
тренировки. У тех животных, у которых рост опухоли при всех
применявшихся воздейст-виях продолжался, преобладающей реакцией был
стресс. Стойкой активации у них вы-звать не удалось. Полученные
данные свидетельствуют о том, что реакцию активации легче поддерживать
на высоких уровнях реактивности низких этажей, т. е. с помощью ма-лой
силы (дозы) воздействия (Гаркави Л. X., Квакина Е.Б. Уколова М. А.,
1977,1979; Гар-кави Л. X., Квакина Е. В., Мулатова А. К., Шихлярова А.
И., 1980; Шихлярова А. П., 1986). Описанный ранее эффект по отношению к
повреждающему влиянию химиолучевой терапии опухолей с помощью вызова
реакции тренировки (при наличии лейкопении) и реакции активации (когда
лейкопения отсутствует) оказался более значительным при ис-пользовании
очень малых по абсолютной величине воздействий, вызывающих эти реак-ции
на высоких уровнях реактивности. Это выражалось в том, что ПеМП меньшей
индук-ции 0,7 мТ, 1,4 мТ, 3,2 мТ) и адреналин в меньших дозах (0,01
мкг) обладали более выра-женным защитным действием (уменьшение
летальности, снижение лейкопении, значи-тельное уменьшение
дистрофических изменений в органах
тимико-лимфатической си-
81
стемы, ткани печени и эндокринных желез) и, вместе с тем, усиливали
противоопухоле-вый эффект при развитии реакции активации. Так,
например, процент торможения роста саркомы-45 при введении сарколизина
(в суммарной дозе 42 мг/кг) увеличивался с 44% до 76% (Квакина Е. Б.,
Гаркави Л. X., Уколова М. А., Тарасьянц Ф. М. Мулатова А. К.,
Гри-горьева Е. Г., 1980; Захарюта Ф. М., 1987, 1989). ПеМП большей
индукции не изменяло противоопухолевого эффекта химиопрепаратов.
То же самое отмечалось и в отношении преодоления признаков
преждевременного старения на уровне организма. Вызов и поддержание в
организме реакции активации, особенно повышенной, с помощью малых по
абсолютной величине доз адреналина, му-мие, малой интенсивности ПеМП
приводило к более быстрому и полному восстановле-нию у старых крыс
полового цикла с появлением в яичниках фолликулов на разных ста-диях
созревания, а у старых самцов отмечалась более выраженная стимуляция
спермато-генеза (более подробно об этом — в главе по старению).
Все это подтверждает представление о том, что реакциям, вызванным
малыми пo абсолютной величине воздействиям, т. е. на высоких уровнях
реактивности, соответствует оптимальный (для данной реакции) комплекс
изменений. Иными словами, физиологической норме соответствует не любая
реакция активации и тренировки, а эти реакции на высоких уровнях
реактивности. При реакциях высоких уровней все подсистемы хорошо
синхронизированы, нет элементов напряженности, свидетельствующих о
десинхрониза-ции работы отдельных подсистем (Гаркави Л. X., Квакина Е.
Б., 1984-1989).
Периодическая закономерность развития
местных адаптационных реакций
Возникает вопрос, существуют ли адаптационные реакции тренировки,
активации и стресса и их периодическое повторение на более низких
иерархических уровнях орга-низма, включая клеточный?
До сих пор говорилось лишь об общих неспецифических адаптационных
реакциях целого организма. А каково их взаимоотношение с местными
— тканевыми, клеточными — реакциями? Явление парабиоза,
открытое Н. Е. Введенским еще в 1904 г. на нервно-мышечном препарате,
представляет собой стереотипно развивающееся нарушение прово-димости
при действии различных по своей природе (качеству, специфике) сильных
раз-дражителей (электрический ток, разные ядовитые и лекарственные
вещества, высокая температура, механическое воздействие) (Введенский Н.
Е., 1951). В дальнейшем Д. Н. Насоновым и В. Я. Александровым (1940)
был описан комплекс
82
неспецифических изменений в клетке, закономерно развивающихся в ответ
на сильное, повреждающее воздействие самых разных по своей природ
факторов. Изучение этих из-менений послужило основой для создания
теории паранекроза, «стремившейся стать основой теории
Введенского о парабиозе, как о своеобразном пограничном состоянии
(между жизнью и смертью) живого организма» (Авцын А. П., Шахламов
В. А,, «Ультраструктур-ные основы патологии клетки». М.,
1979. С. 35). Последователи Г. Селье говорят о местном стрессе
(Календо Г. С, 1972). При местном стрессе также имеются черты
поврежде-ния, как и при общем — элементы раздражения и
дегенерации клетки. Г. С. Календо (1972—1982), помимо клеточного
стресса, описала другой неспецифический комплекс изменений в ответ на
действие раздражителей небольшой интенсивности. Есть ли местная реакция
активаций и местная реакция тренировки? Когда в организме отсутствует
какой-либо патологический процесс или очаг повреждения, местная реакция
является частью общей. В нормальном здоровом организме местные реакции
подчинены регулирующим влияниям общих. Поэтому морфологические
изменения (на тканевом и клеточном уровне) органов тимико-лимфатической
системы, эндокринных желез, нервных структур мозга, характеризующие
реакции тренировки, активации (спокойной и повышенной) и стресса,
являются признаками и соответствующих местных
реакций.
М. А. Шишло полагал, что фундаментальным надмолекулярным механизмом
раз-вития адаптационных реакций организма является изменение зарядовой
асимметрии мем-бран, через которое осуществляется регуляция
автоколебательных процессов в живых структурах (Холодов Ю. А., Шишло М.
А., 1979).
Изучение биофизических показателей функциональной активности мембраны
при различных адаптационных реакциях обнаружило при стрессе резкое
повышение транс-мембранного потенциала, микровязкости, увеличение
восстановленности сульфгидриль-ных групп. Величины этих показателей
снижались по мере чередования реакций в после-довательности: стресс
— тренировка — активация — переактивация (Курышев И.
Е., 1987). Таким образом, И. Е. Курышев также приходит к выводу о
реализации гомеостати-ческих механизмов в
сложных системах через
биоритмические процессы в мем-бране.
Изучение мембрано-рецепторного
аппарата тимоцитов и синтетической активности тимоцитов и лимфоцитов
периферической крови (Шварцбурд П. Н., 1980) также выявило
(четкую корреляцию этих показателей с характером адаптационной
реакции; наиболее развитый мембрано-рецепторный аппарат и высокая
синтетическая активность клеток (по соотношению
односпиральных и двуспиральных нуклеиновых кислот) наблюдались при
реакции активации (особенно
83
повышенной), а слабо развитый мембрано-рецепторный аппарат и наиболее
низкая синте-тическая активность — при стрессе.
Синтети- ческая активность лимфоцитов перифери-ческой крови
при изменении адаптационной реакции изменяется даже
несколько раньше,
чем формула белой крови.
В щитовидной железе отмечалось снижение синтетической активности
тиреоцитов при стрессе и повышение — при реакции активации
(Гаркави Л. X., Квакина Е. Б., Шейко Е. А., 1987). С
появлением какого-либо местного патологического
процесса (опухоли, воспаления, повреждения и т. д.) стимулируются
древние автономные механизмы саморе-гуляции клеток, автономность
клеточных реакций резко возрастает (Крыжановский Г. Н., 1973;
Гаркави Л. X., Квакина Е. Б., 1984). Клетка двойственна по своей
природе. В про-цессе эволюции клетка вначале существовала в виде
самостоятельного организма, а затем стала субъединицей все более
сложных многоклеточных организмов с многоуровневой иерархией. В
этом новом качестве клетка подчиняется регулирующим влияниям организма,
но вместе с тем сохраняет и саморегуляцию, черты автономности. В
условиях па-тологии уже не только реакции организма подчиняют своему
влиянию местные реакции, но и местные реакции могут подчинить своему
влиянию общие, навязать организму свой характер изменений. При
воспалительных процессах местный стресс, развивающийся в очаге
воспаления, может быть либо подавлен, либо .
навязывает свою реакцию целому организму. Это одна из причин
того, что хотя
общий стресс и
обладает про-воспалительным действием,
но полностью излечить
воспаление на фоне общего стресса,
угнетающего защитные системы организма, крайне трудно— в
большинстве слу-чаев происходит переход воспаления в хроническую вяло
текущую форму. В то же время, при развитии реакции
активации, хотя и
обладающей провоспалительным действи-ем,
воспаление течет более быстро и завершается полным выздоровлением
благодаря высокой функциональной
активности защитных систем. Но может быть и наоборот: обширная
воспалительная реакция навязывает стресс всему организму, преодолевая
об-щую реакцию активации, недостаточно стойкую в этом случае.
Если о признаках общих неспецифических адаптационных реакций мы судим
по изменениям тимико-лимфатической системы,, эндокринных желез и
центральной нервной системы, то о признаках разной тканевой или
клеточной реакции можно судить по тем изме-, нениям, которые
характеризуют этот иерархический уровень организма. Можно сказать, в
самом общем виде, что при активации отмечаются признаки
физиологической, умеренной стимуляции, при стрессе — признаки
либо угнетения и дегенерации, либо ги-перстимуляции. При реакции
тренировки нет ни выраженных признаков стимуляции, ни угнетения и
дегенерации. Эти общие положения
84
имеют конкретное выражение и в морфологических, и в гистохимических
характеристи-ках
клеток.
Возникает также вопрос: наблюдается ли на тканевом уровне
та периодическая закономерность развития неспецифических адаптационных
реакций, которая была найде-на на уровне организма Происходит ли на
клеточном уровне — по мере увеличения силы (дозы)
воздействия — повторение тетрад адаптационных реакций:
тренировки, спокой-ной активации, повышенной активации и стресса? Были
проведены опыты по влиянию переменного магнитного поля напряженностью
0,1 и 1 мТ в разных «дозах» (по
дли-тельности экспозиции и частоте) на культуры
клеток (АМН, Нер-2, ПЧ и ФЭЧ). Изу-чалось
количество погибших клеток и, таким образом, оценивалась степень
защитного эффекта магнитного поля. «Доза» менялась по
экспозиции последовательно с использо-ванием коэффициента общих
адаптационных реакций. В результате были отмечены вол-нообразные
колебания числа погибших клеток с высокой степенью достоверности
разли-чий (Гаркави Л. X., Квакина Е. Б., Барсукова Л. П.,
Шихлярова А. И., Берман М. В., Ку-рышев И. Е., 1982). Аналогичную
зависимость удалось обнаружить при определении
чувствительности опухолевых
клеток (экспериментальных опухолей крыс: карцино-мы
Герена, РМК-1, саркомы-45 и опухолей яичника человека) к разным дозам
алкили-рующих химиопрепаратов. Чувствительность определялась
по ингибированию сульф-гидриль-ных групп (Кулик Г. И., 1964).
Наблюдалось волнообразное изменение чувстви-тельности в зависимости от
дозы препарата (тиоТЭФ, сарколизин): при
относительно большой, терапевтической дозе — снижение
уровня SH-групп (ингибирование), местный стресс; при уменьшении
дозы в 1,25 (в 1,5) раза — наиболее высокий уровень
SH-групп, местная реакция активации; при последующем таком же
уменьшении — промежуточный уровень,
местная реакция трени-ровки. При дальнейшем
последовательном уменьше-нии терапевтической дозы препарата с таким же
шагом закономерность развития местных реакций в зависимости от
относительной величины дозы повторялась многократно (Гар-кави Л.
X., Квакина Е. Б., Мулато- А. К., Тарасьянц Ф. М., 1981; Захарюта Ф.
М., 1989). Та же зависимость сохранялась при определении
чувствительности (но этому же показате-лю) не только к
алкилирующим препаратам, но и к разовым дозам гормона (синестрола) и не
только в клетках опухоли, но и в нормальных тканях (печень
и эритроциты), что гово-рит о ее специфическом характере.
Если предположить, что в противоопухолевом действии
химио-препаратов суще-ственную роль играют неспецифические клеточные
реакции, становится ясным, что для угнетения клеток опухоли нужно
вызывать местный стресс. Сочетание местного стресса с реакцией
активации организма должно привести к наилучшему противоопухо-
85
левому эффекту. Применение доз химиопрепаратов, в 5 раз меньших
терапевтической (на более высоком уровне реактивности), но к которым
опухоль была чувствительной так же, как к терапевтической, показало
в эксперименте наличие такого же
противоопухолево-го эффекта, как и от терапевтической дозы. Состояние
животных, конечно, было значи-тельно лучшим. В результате
морфологических исследований выяснилось, что при этом не отмечалось
выраженного (как при использовании
терапевтической дозы химиопре-парата) угнетения
лимфоидной ткани органов тимико-лимфатической
системы и таких эндокринных желез, как щитовидная железа, половые
клетки, клубочковая зона
коры надпочечников (секретирующая
минералокортикоиды). Пучковая зона коры надпочеч-ников также была
в состоянии высокой, но не чрезмерной, функциональной активности. Эти
данные говорят в пользу существенной роли в противоопухолевом действии
алкили-рующих химиопрепаратов неспецифических местных адаптационных
реакций, а также свидетельствуют в пользу существования на уровне
тканей и клеток периодической зако-номерности повторения
основных адаптационных реакций по мере увеличения силы
(дозы) воздействия (Гаркави Л. X., Квакина Е. Б., Шихлярова А. И.,
Тарасьянц Ф. М., 1982; Гаркави Л. X., Квакина Е. Б., 1984; Захарюта Ф.
М., 1989).
Таким образом, было показано, что и местные адаптационные реакции
подчинены принципу периодической зависимости от количества действующего
фактора, а не только от его качества, специфики, т. е. и на уровне
тканей и клеток существуют и влияют на ме-стные процессы
неспецифические адаптационные реакции. Сочетание общих и местных
реакций открывает хорошие перспективы для дальнейших исследований по
управлению состоянием организма и борьбы с патологическими
процессами и повреждениями, имеющими не только общий, но и местный
характер.
Сочетание неспецифических и специфических реакций
А как обстоит дело с сочетанием неспецифических и специфических
реакций? Ведь никто не отрицает наличия специфического действия
различных раздражителей. Ка-ждый раздражитель вызывает какую-либо
неспецифическую реакцию и одновременно ка-кие-то свои, свойственные
данному действующему фактору, специфические изменения, которые
накладываются на неспецифическую основу. Поэтому и стресс, и активация,
и тренировка имеют, кроме основных черт сходства, и какие-то отличия,
связанные со спе-цифическим действием фактора, который вызвал эти
реакции. Если связать изменение эффективности с силой (дозой)
воздействия, можно сказать, что для неспецифических ре-акций это
происходит нелинейно, дискретно: повышение эффективности
сменяется сни-жением в небольшом
86
диапазоне изменения силы. Вместе с тем специфическая эффективность
воздействия (расширение коронарных сосудов, снижение артериального
давления, снижение коагуля-ционных свойств крови и т. п.)
сохраняется линейно в довольно большом диапазоне — в пределах
терапевтических доз. Такую линейность специфического действия можно
свя-зать с тем, что влияние происходит на уровне эффекторного звена
(точка приложения),': через местные реакции. Кроме того, большое
значение, по нашему мнению, имеет струк-турно-функциональная
специализация (органа, ткани, клетки), которая подразумевает на-личие
линейной и однозначной зависимости эффекта от силы воздействия
(например, от-вет мышечной ткани на увеличение нагрузки). Мы считаем,
что соединение оптимального неспецифического эффекта со специфическим
при возможной минимизации дозы должно быть наиболее эффективным. При
таком сочетании получено, как уже говорилось, повы-шение
противоопухолевой эффективности алкилирующих химиопрепаратов. При
лечении психических заболеваний реакцию активации вызывали
психотропными средствами (Ше-велев А. И., Невский М. П., Гаркави Л. X.,
Уколова М. А., Квакина Е. Б., 1981), при лече-нии некоторых
сердечнососудистых заболеваний — спазмолитическими,
гипотензивными и т. п. Результаты исследований показали, что доза
средств специфического действия на фоне реакции активации может быть
значительно уменьшена по сравнению со средней терапевтической без
снижения специфического эффекта и с улучшением общего состоя-ния
организма.
Таким образом, можно сказать, что оптимальный эффект достигается
тогда, когда нужное специфическое
действие (например, расширение коронарных сосудов или
по-вреждение клеток опухоли) сочетается с повышением резистентности
организма, с физио-логической гармоничной
стимуляцией функциональной
активности его основных систем, т. е. с развитием
адаптационной реакции активации высоких уровней реактивно-сти. Если тe
же специфические средства одновременно вызывают стресс, т. е.
выраженное в разной степени угнетение защитной тимико-лимфатической
системы и дисгармонии в работе нейроэндокринной системы,
неспецифическое действие будет снижать эффектив-ность специфического.
Кроме того, часто специфическое можно характеризовать как оп-ределенное
количество неспецифического. Так, например, специфическим
действием ад-реналина считается повышение количества сахара в
крови и артериального давления. Это действительно так при использовании
больших, чаще всего стрессорных доз адреналина. В небольших
дозах, вызывающих реакцию активации
высоких уровней реактивности, эффект адреналина
противоположный. Нужно помнить, что количество переходит в каче-ство, и
это также обусловливает специфическое действие.
Разная степень повышения или снижения какого-либо
87
показателя тоже может свидетельствовать о качественно отличных состояниях.
Естественно возникает вопрос о сознательном целенаправленном сочетании
спе-цифического и неспецифического действия применяемых лечебных
средств и воздейст-вий. Это вполне реально, так как специфическое
средство сохраняет свое специфическое действие в довольно широком
диапазоне дозы (силы), а переход из одной неспецифиче-ской
адаптационной реакции в другую происходит уже при изменении дозы чаще
всего на 20%. Поэтому, специально подбирая дозу (силу) специфического
воздействия под контро-лем простых показателей неспецифических
адаптационных реакций, можно существенно повысить эффективность лечения.
Таким образом, наличие и неспецифических, и специфических реакций,
понимание сходства и отличия закономерности их развития открывает
перспективу правильного со-четания этих реакций.
Синхронизация, резонансные явления, энтропия, негэнтропия в периодиче-ской системе адаптационных реакций
Каким образом осуществляется связь между местными и общими
реакциями, т. е. связь между работой различных
иерархических уровней организма, вплоть до клеточно-го, и организмом в
целом как системой? По мнению В. П. Казначеева и Л. П. Михайловой
(1985), объединению организма в систему способствуют электромагнитные
излучения. Это не противоречит представлениям о роли
синхронизации колебательных процессов, имеющих
различные характеристики на различных иерархических уровнях. Была
выска-зана гипотеза (Л. X. Гаркави) о
том, что каждой адаптационной реакции на разных ие-рархических уровнях
организма присущи свои частоты, которые подстраиваются к более
длительным ритмам. Известно, что в процессе действия любого
раздражителя в организме происходит многократное перекодирование
частоты в силу и наоборот. При этом частот-ные характеристики реакций
должны быть общими для самых различных живых организ-мов,
сформированных в процессе эволюции под влиянием одних и тех же
экзогенных час-тот. Иными словами, индивидуальные (а возможно, и
видовые) различия связаны с опре-деленным коэффициентом
пропорциональности (К) перекодирования силы в частоту.
Для проверки этого предположения были проведены исследования действия
различных частот переменного магнитного поля на мозг крыс (Барсукова Л.
П., Гаркави Л. X., Котля-ревская Е. С, Марьяновская Г. Я., 1983).
Выяснилось, что увеличение частоты действует аналогична увеличению
интенсивности (Гаркави Л. X., Квакина Е. Б., 1984): приводит у
большинства животных (кроме
88
тех, кто находился в стойких состояниях к периодическому чередованию
адаптационных реакций тренировки, спокойной активации, повышенной
активации и стресса. При этом коэффициент реакции оказался меньше, чем
при варьировании в силе (дозе). Было показано, что некоторые частоты
при однократном воздействии вызывают преимущественно развитие реакции
активации, некоторые — стресса, и обнаружена частота, в
большинстве случаев не вызывающая изменений. Однако повторение
воздействий довольно быстро (в течение недели) приводило либо к срыву в
стресс, либо к переходу в напряженные реак-ции. По-видимому, это
связано с применением одной частоты, вызывающей резонансные явления
лишь на одном из иерархических уровней, тогда
как обычно в организме од-новременно (или
последовательно) работает целый набор частотных характеристик на разных
иерархических уровнях, что приводит к смягчению резонансных явлений. В
на-стоящее время нами используется либо аккорд частот (одновременное
использование не менее трех частот, свойственных
разным иерархическим уровням
организма), либо арпеджио — применение нескольких
частот последовательно (Гаркави Л. X., Квакина Е. Б.,
1987), от более низких, свойственных ЦНС, как регулирующей системе
организма в целом, до более высоких, присущих отдельным подсистемам,
тканям, клеткам.
Синхронизацию считают основным системообразующим фактором (Путилов А.
А., 1987). Степень синхронизации различна при разных реакциях. Хорошо
известна десин-хронизация при стрессе, которая наиболее наглядна при
стрессе, вызванном быстрой или многократной сменой часовых поясов
(Моисеева Н. И., Сысуев В. М., 1981; Питтендрих К., 1984; и др.).
Известно, что десинхронизация в организме наблюдается при старении
(Aschoff J., Gereche U., Wever R., 1967; Pittendrigh С S., Daan S.,
1974; Mohan C, Radha E., 1978; Ehret C. F., Groh K. R., Meinert J. C.,
1978; Halberg F., Nelson W., 1978; Wever R., 1975; Дэвис Ф., 1984).
Напротив, оптимальная синхронизация свойственна реакции акти-вации,
особенно повышенной активации высоких уровней реактивности. Это
понятно, по-скольку синхронизация считается «энергетически
оптимальным режимом функциониро-вания динамической системы и
обеспечивает ей свойства целостности и устойчивости» (Путилов А.
А. Системообразующая функция синхронизации в живой природе.
Новоси-бирск, 1987. с. 31).
Уже говорилось о том, что на низких уровнях
реактивности (высоких этажах) реак-ции тренировки и активации теряют
присущую им синхронизацию, появляются признака десинхронизации,
которые можно выявить даже при морфологическом исследовании
бе-лой крови. При стрессе, особенно низких уровней реактивности,
наряду с десинхрониза-цией отмечается гиперсинхронизация отдельных
процессов, что обусловливает, например, гиперстимуляцию системы
89
АКТГ — глюкокортикоиды. Развитие местного патологического
процесса, особенно его хроническое течение, почти всегда сопровождается
десинхронизацией местной и общей
неспецифической редакции.
Тенденция к синхронизации есть, по существу, тенденция к
ритмичности, упоря-доченности, гармонии (Блехман И. И., 1971, 1981),
противостоящая тенденциям в
при-роде, постулируемым вторым законом,
термодинамики. В отличие от неживых систем, в которых, согласно второму
началу термодинамики, происходит лишь снижение
упоря-доченности и, соответственно,
рост энтропии, приводящей к необратимому
равно-весию, в процессе развития живых систем энтропия
может не только расти, но и умень-шаться, т. е. они способны к
производству негэнтропии. Согласно Э. С. Бауэру (1935), для живых
систем характерно состояние устойчивого неравновесия при котором
постоянно совершается работа против равновесия и строятся
специфические для живого
нерав-новесные структуры, обладающие так называемой
свободной, или структурной, энерги-ей. Это и обеспечивает сохранение
неравновесности живой системы в условиях
«вырав-нивающего» влияния окружающей среды. В
процессе жизнедеятельности
происходит повышение энергетической и
структурной упорядоченности живой системы, что свиде-тельствует о
снижении энтропии и увеличении негэнтропии. Таким образом, поведение
живых систем не укладывается в рамки классической термодинамики —
термодинамики линейных
необратимых процессов, протекающих вблизи от
состояния термодинами-ческого равновесия. И.
Пригожиным (1960) сформирован
принцип «упорядоченность через флуктуации» и
совместно с П. Глансдорфом (1973) выдвинута теория развития от-крытия
систем на основе термодинамики необратимых нелинейных процессов.
Таким образом, живой организм никогда не находится в равновесии.
Равновесие для живого означает смерть. Какие же состояния увеличивают
неравномерность живых систем, негэнтропийную тенденцию живого? Согласно
И. А. Аршавскому (1982, 1986), сюда относится избыточно-анаболическая
фаза «физиологического стресса», в процессе которой
происходит избыточное образование «живой протоплазменной
массы» (пролифе-рация и гиперплазия, гипертрофия) и свободной
структурной энергии, обогащение энер-гетического фонда, возрастание
физических и интеллектуальных возможностей организ-ма. Напротив,
катаболическая фаза, по Аршавскому, когда происходит деструкция,
отли-чается максимальным производством энтропии.
Если рассмотреть в плане энтропийно-негэнтропийных процессов
адаптационные реакции организма, то, исходя из характеристики различных
реакций на разных уровнях реактивности, можно заключить,
что наибольшей неравномерностью и
максимумом нег-
90
энтропии (и, соответственно, минимумом энтропии) обладает реакция
повышенной акти-вации высоких уровней, а наиболее близким к
термодинамически равновесному состоя-нию с минимумом негэнтропии (и,
соответственно, максимумам энтропии) — реакция стресс низких
уровней реактивности. В целом, при одноименных адаптационных реакциях
реакции высоких уровней характеризуются большей неравновесностью и
негэнтропией. В этом можно убедиться, анализируя под соответствующим
углом зрения изменения в орга-низме при разных адаптационных реакциях
(от гиперплазии лимфоидной ткани и увели-чения синтетических процессов
в эндокринных органах при реакции активации, особенно] повышенной
активации высоких уровней реактивности, в отличие от процессов
гипопла-зии лимфоидной ткани, снижении синтетических процессов в
эндокринных, органах при стрессе, особенно низких уровней,— до
новообразования синаптических окончаний в центральной нервной системе и
периферических нервных образованиях при активации высоких уровней
и дегенерации их при стрессе низких и т. п.).
Таким образом, негэнтропийные и энтропийные тенденции в развитии
организма по-разному выражены при различных реакциях и на
разных уровнях реактивности. От-сюда следуют важные практические
выводы, которые еще будут сделаны при рассмотре-нии проблемы
старения, резервов организма и вопросов активационной терапии на
осно-ве теории адаптационных
реакций.
С резонансными и
синхронизационными явлениями связана
дискретность адаптационных реакций. Если каждой реакции свойственны
определенные частотные ха-рактеристики, с помощью которых они могут
быть вызваны, резонансные, явления долж-ны возникать периодически, по
мере повторения этих и кратных им частот. Отсюда выте-кает принцип
дискретности в адаптационной деятельности организма. Само по себе
нали-чие разных типов адаптационных реакций и их закономерное
повторение при увеличении действующего фактора на определенную
величину (коэффициент реакции) уже говорит о важнейшей роли
резонансных явлений в их развитии, подобно тому, например, как
обу-словлена резонансами дискретность орбит
«эволюционно-зрелых» динамических систем (Молчанов А.
М.,
1966).
Дискретность (квантованность) состояний,
скачкообразность перехода из одной реакции в другую наблюдается и на
примере различных показателей. Так, процентное со-держание лимфоцитов
при переходе из тренировки в активацию возрастает (можно ду-мать
о постепенном переходе), а при переходе из активации в стресс
— резко падает (скачок). Четкое разделение реакций по
количеству лимфоцитов удалось произвести и потому, что
градуальному увеличению числа лимфоцитов от тренировки до активации
со-ответствует дискретное изменение показателей, линейность
которых нарушается
91
в области пограничных для этих реакций значений числа лимфоцитов (см.
рис. 38). Так, секреция глюкокортикоидов при переходе из тренировки в
активацию снижается, а при переходе из активации в стресс резко
нарастает.
Дискретность состояний также выражена при переходе с одного уровня
реактивно-сти на другой, так как это либо смена стресса на тренировку
(при увеличении этажа), либо наоборот (при снижении этажа). Различия
между этими состояниями — и по комплексу изменений в основных
системах организма, и по энергетике, и по уровню резистентности
настолько значительны, что можно говорить о резком скачке.
Принципу дискретности реакций как принципу, увеличивающему
адаптационные возможности организма, придается большое значение и с
философских позиций (Щепин О. К., Царегородцев Г. И., Ерохин В. Г.,
1983).
Представление о дискретности реакций позволяет понять, почему живой
организм может реагировать не на сумму всех раздражителей, а на
какой-то раздражитель отдельно, выби-рая его из массы падающих
раздражителей. Так, при стойкой повышенной активации вы-соких уровней
реактивности организм приобретает способность выбирать малый по
абсо-лютной величине раздражитель, соответствующий тому уровню, на
котором поддержива-ется реакция организма. При длительном стрессе и
переактивации низких уровней реак-тивности организм теряет эту
способность и может выбирать лишь большие по абсолют-ной величине
раздражителя, также соответствующие уровню реактивности, на котором он
длительно функционирует. Таким образом, на уровне общих реакций
организма осуществляется положение И. И. Шмальгаузена (1968) о том, что
каналы связи биологических систем с внешней средой контролируются и
регулируются самой системой в зависимости от ее состояния.
Способность к активному выбору определенной реакции так же, как
целенаправ-ленный вызов реакции, реализуется благодаря резонансному
взаимодействию: частота внешнего фактора (так называемого
«принудителя» ) должна быть близкой или краткой частоте
«колебателя», свойственной данной реакции. Это
соответствует известному по-ложению о том, что в нелинейных системах
нарушается принцип суперпозиции, и реак-ции их зависят не от суммы
падающих воздействий, а от присущих им внутренних свойств.
Дискретность реакций и преимущественная роль слабых (информационных)
взаи-модействий объясняет принципиальную возможность получения реакций
на слабые раз-дражители в присутствии влияния сильных, т. е., например,
перевод опухолевого стресса низких уровней реактивности в реакции
тренировки или активации более высоких уров-ней. Подтверждение такой
возможности имеется и в некоторых физических законах, на-пример, в
принципе наименьшего
92
действия. Так, при стрессе (особенно высокоэтажном) наиболее велики
энергетические траты, и если на организм падает раздражитель, на
который он может ответить с меньши-ми тратами, то это для него более
легкий и поэтому более приемлемый путь. Кроме того, из синергетики
известно, что для воздействия на сложную открытую систему необходимо
учитывать законы, но которым она работает, чтобы перевести систему в
одно из ее устой-чивых состояний (наша цель — в состояние
повышенной активации высоких уровней). «Бесполезно тратить
энергию и время на насилие над сложными системами. Надо знать, как они
функционируют, и с минимальными усилиями возбуждать то, что им
адекватно. Нужно учитывать собственные реакции системы на внешние
воздействия» (Курдюмов С. П.,
1988).
Изучение реакций на разных уровнях реактивности показало, что только
реакции тренировки, и особенно активации низких этажей могут считаться
физиологическими и претендовать на связь с понятием «идеальная
норма» (по В. М. Дильману). Адаптационные реакции низких
уровней реактивности — тренировки и активации, наряду со
стрес-сом разных уровней реактивности, являются, по нашему
мнению, неспецифическими со-стояниями, реализующими с помощью
положительной обратной связи закон отклонения гомеостаза, приводящий к
развитию «нормальных» болезней и старения (Дильман В.
М„ 1968-1987).
Наличие сложной периодической зависимости типа адаптационной реакции от
ко-личества действующего фактора существенно расширяет адаптационные
возможности ор-ганизма, позволяет реагировать на тонкие изменения
наиболее экономичным способом. Очень важно, что каждая реакция
формируется под влиянием одновременно двух состав-ляющих: абсолютной
величины раздражителя, под влиянием которой осуществляется вы-бор
уровня реактивности, и относительной величины, определяющей характер
реакции на этом уровне.
Знание этой закономерности открывает большие перспективы для тонкого
управ-ления состоянием организма с целью его
оптимизации.
Система ареактивности в поддержании гомеостаза
В процессе эволюции, кроме принципа реагирования, сложился и другой
принцип адаптации — принцип ареактивности, который, на первый
взгляд, кажется более далеким- от состояния устойчивого неравновесия,
характерного для живого.
При изучении периодической системы адаптационных реакций обнаружилось,
что часто после стресса любого уровня выявляется своеобразная зона,
названная зоной ареак-тивности, в которой види-
93
мая реакция отсутствует (Гаркави Л. X., Квакина Е. Б., 1975). Понятно,
что раздражитель меньше слабого, тренировочного не вызывает никаких
изменений, являясь как бы подпо-роговым. Но как понять, что
раздражитель больший, чем стрессорный для данного уров-ня, не вызывает
никаких сколько-нибудь заметных изменений ни в соотношении формен-ных
элементов белой крови, ни в функциональной активности щитовидной железы
или коры надпочечников (по исследованным нами показателям)? Отсутствие
видимой реакции на раздражитель отмечалось и ранее. В опытах Н. П.
Кравкова (1924) и И. П. Чукичева (1958) кровеносные сосуды уха кролика
не реагировали на определенные дозы некоторых веществ, в то время как и
увеличение и уменьшение дозы оказывало действие. Опреде-ленный диапазон
раздражителей, в котором отсутствовала реакция, Кравков назвал
«ней-тральной зоной». На подобное явление особое внимание
обратил А. И. Венчиков
(1962), назвав его «зоной
бездействия вещества». С фактом отсутствия реакции на ударные
(большие, чем терапевтические) дозы противоопухолевых токсических
препара-тов столкнулись М. А. Уколова и Г. А. Веревкина (1971). При
введений тиоТЭФа в так называемой терапевтической дозе (2 мг/кг)
развивался комплекс изменений,
типичный для стресса. Применение
значительно больших доз (7,5 мг/кг и 15 мг/кг) приводило часть
животных (50%) к гибели с наличием резких изменений,
характерных для стресса (уменьшением почти втрое веса тимуса и
селезенки и увеличением почти вдвое веса надпочечников). В
то же время у выживших животных существенных изменений в
эндок-ринной системе (при использовании
тех же методов исследований) и морфологиче-ском
составе белой крови не наблюдалось. Эту своеобразную реакцию Уколова
назвала «реакцией блокады», считая, что действие
больших доз тиоТЭФа блокируется в ЦНС. Л. Д. Кисловский
(1984) считает, что наличие
«зон ареактивности» (зон бездействия)
связано с включением в работу компенсирующих «защитных»
механизмов.
Исследования показали, что зона ареактивности часто повторяется между
всеми триадами адаптационных реакций, когда абсолютная величина (сила,
доза) раздражителя очень мала и когда она очень велика, даже близка к
летальной. Возможно, адаптация в смысле «привыкания»
объясняется развитием «зоны ареактивности» как при
увеличении или уменьшении силы раздражителя (до определенной величины),
так и при соответст-вующем уменьшении или увеличении чувствительности.
Когда отмечается стрессорная ареактивность, то по показателям
лейкоцитарной формулы наблюдается типичный стресс, а изменения в
тимико-лимфатической и эндокринной системах мягче, чем при реакции
стресса, вызванного той же по абсолютной величине дозой раздражителя.
Характерно, что при ареактивности показатели белой крови в динамике не
претерпевают
94
никаких изменений. Вывести из стрессорной ареактивности труднее, хотя
это состояние менее повреждает организм, чем реакция хронического
стресса. Особенно часто встреча-ется стрессорная ареактивность на
низких уровнях реактивности (высоких этажах) при длительном,
систематическом действии большого по абсолютной величине
раздражителя.
Не только нами, но и другими исследователями было отмечено, что
стрессорные воздействия, первоначально вызывавшие соответствующую
реакцию гипофиз-адренало- вой системы (например, подъем концентрации
кортикостерона в крови или надпочечни-ках), после многократного
повторения переставали оказывать видимое влияние, хотя при-сущий
стрессу гормональный фон в организме сохранялся (Mikulaj L. et al.,
1974; Виру A. A., 1974; Keim К., Sigg Е., 1974; Меерсон Ф. 3.,
Пшенникова М. Г., 1988). Состояние акти-вационной ареактивности,
напротив, отмечается на высоких уровнях реактивности, при длительном
поддержании в организме повышенной активации. По своим характеристикам
это еще более благоприятное состояние, чем соответствующая реакция, и.
обладает боль-шей стойкостью. Состояние тренировочной ареактивности
встречается крайне
редко.
Близким к понятию ареактивности является переактивация —
напряженная по лимфоцитам (лимфоцитов) реакция повышенной активации
низких уровней реактивности.
Таким образом, состояния ареактивности отличаются:
1. Отсутствием заметной реакции в большом диапазоне величины раздражителя.
2. Стойкостью.
3. Наличием более благоприятных изменений в организме, чем при
соответствую-щей реакции, вызванной тем же по абсолютной величине
раздражителем.
С чем может быть связана природа ареактивности? Мы подошли к
осмысливанию состояния ареактивности с позиций представлений об
организме как о сложной колеба-тельной системе.
Известно, что стойкость колебательный процессов возрастает с
увеличением часто-ты колебаний. Поэтому было высказано предположение,
что состояние ареактивности включает в себя как частотные
характеристики основных реакций (Л. Х. Гаркави), так и колебания,
возбуждаемые параметрически (Е. Б. Квакина) с помощью так называемого
«параметрического резонанса». Такие колебания возникают в
результате изменения (по разным причинам) «колебательных
параметров» системы (в данном случае — организма),
вследствие чего многие из этих параметров начинают резонировать с
различными внеш-ними воздействиями. Существенно, что исходные колебания
могут быть сколь угодно ма-лы, но благодаря интенсивной энергетической
подпитке, например, дважды за один пери-од колебаний (при соотношении
собственных и входных частот как 1:2), происходит их быстрая раскачка.
В системе начинают совершаться
95
нарастающие (некоторое время) колебания около положения равновесия,
способствуя поддержанию ее устойчивого неравновесного состояния —
основного, по Бауэру, свойст-ва живых систем. В формировании реакций
участвует лишь часть структурных элементов систем, органов, тканей. Не
участвующий в реакции резерв структурных элементов нахо-дится в
состоянии ареактивности. Поскольку в такой сложной колебательной
системе, как организм, всегда найдется множество малых собственных
колебаний, которые будут сов-падать по фазе с внешними, возникает почти
непрерывный спектр колебаний и, соответ-ственно, возможность
энергетической подпитки от множества внешних факторов, среди которых
большое место занимают физические полевые факторы, о чем
свидетельствуют исследования А. П. Дуброва (1969), В. П. Казначеева
(1985) и др. Это обусловливает большую энергетическую выгодность
состояний ареактивности по сравнению с реакцией соответствующего
уровня. Энергетическое преимущество, возможно, является одним из
факторов, обусловливающих высокую устойчивость состояний ареактивности.
Поэтому даже патологические стойкие состояния, связанные с
ареактивностью, более благоприят-ны для организма, чем соответствующая
реакция на воздействие той же силы, в том числе, и состояние
ареактивности при росте опухоли, которое является все же мерой защиты,
хо-тя и крайней. По-видимому, для формирования какой-либо реакции
необходимо участие определенного минимума структурных элементов. Если
этот минимум не достигнут, раз-вивается состояние ареактивности.
Высказывается предположение (Л. X. Гаркави), что ареактивность —
это фоновое живое состояние, осуществляющее энергетическое обога-щение
организма резонансным путем, что соответствует представлениям о
накопительной функции «покоя» — отсутствия реакций,
приводящих к трате энергии. Однако это со-стояние покоя является весьма
относительным, поскольку по частоте колебательных про-цессов
ареактивность превышает реакции, так как включает в себя их частотные
состав-ляющие. Вместе с тем наличие такого фона энергетически облегчает
развитие любой ре-акции, так как в фоновом состоянии уже имеются
свойственные каждой реакции частот-ные характеристики, позволяющие
использовать резонансный путь для развития реакции.
Экспериментальные и клинические исследования показали, что изменения
неспецифиче-ских адаптационных реакций и соответствующих им состояний
ареактивности по мере увеличения или уменьшения абсолютной величины
действующих факторов подчиняются разным количественным закономерностям.
Спонтанное развитие состояний ареактивно-сти происходит, чаще всего,
лишь при длительном поддержании в организме одной и той же реакции
(роль повторности
воздействия).
Все сказанное заставило нас отказаться от термина «зона ареактивности», заменив его
96
«состоянием ареактивности». Комплекс различных
состояний ареактивности, подчи-няющийся своей
количественно-качественной закономерности, составляет систему
аре-активности, диалектично
дополняющую систему реакций.
Наиболее адекватно су-ществующие количественные отношения можно
представить в виде двойной спирали (предложена Л. X. Гаркави), причем
спирали расположены коаксиально и расширяются кверху (а возможно, и
книзу) по мере увеличения (а возможно, и уменьшения) количества
воздействий от некоторой нулевой величины. Адаптационные реакции
описываются внутренней спиралью, а состояния ареактивности —
внешней (рис.41).
Рис. 41. Трехмерная диаграмма динамики реализации периодической системы
адап-тационных реакций и ареактивности: z — основная си ловая
ось, соответствующая значениям величины действующего фактора для
реакций и уровней реактивности. В плоскости х, у отображается
адаптационная реакция, на оси х и у могут быть отло-жены 2 любых
сопряженных показателя адаптационных реакций.
Использование образа двух спиралей
иллюстрирует наличие двух типов адаптаци-онных процессов: адаптационных
реакций и соответствующих реакциям состояний ареак-тивности. При
обработке на ЭВМ ЕС-1037 большого массива данных по результатам
ле-чения больных с двигательными нарушениями центрального генеза с
помощью неспеци-фических адаптационных реакций между типом ре акций и
эффективностью лечения была обнаружена зависимость в виде двойной
спирали. Такая трехмерная диаграмма может быть применена как для
теоретического, так и для экспериментального анализа процессов
гомеостаза (Гаркави Л X., Квакина Б. В., Серегин Ю. В., Файдыш Е. А.,
1985). Эта диа-грамма позволяет понять с позиций колебательных
процессов «перескоки» с витка на ви-ток по каждой спирали и
с одной спирали на другую со скачкообразным изменением ре-зонансной
частоты. При этом становится наглядной роль амплитуды: при длительной
по-вышенной активации высоких
97
уровней реактивности максимально возрастает синхронизация колебаний, и
увеличение амплитуды способствует «перескоку» на спираль
ареактивности. Гиперсинхронизация при переактивации низких уровней
реактивности также способствует переходу на спираль ареактивности. При
стрессе, особенно низких уровней реактивности, как уже говорилось,
происходит десинхронизация большинства процессов и гиперсинхронизация
— меньшин-ства. В результате образуются более медленные волны с
очень высокой амплитудой, что способствует переходу на спираль
ареактивности. Эта диаграмма отражает представление о разных типах
резистентности, обнаруженных в эксперименте и клинических
исследова-ниях. Кроме описанной ранее активной и пассивной
резистентности, обусловленной ти-пом реакции, имеются два вида
резистентности, связанные, как и большинство стойких состоянии, с
ареактивностью. Один из них связан с состоянием ареактивности
повышен-ной активации высоких уровней реактивности (низких этажей). Это
«подвижный» тип ре-зистентности, основанный на
использовании принципа устойчивого неравновесия Э. Бау-эра, который
характеризуется частыми переходами с одной спирали на другую. Такой тип
резистентности лежит в основе устойчивого здоровья. Второй тип
резистентности связан с состоянием стресса и напряженной повышенной
активации (переактивации) низких уров-ней реактивности (высокие этажи).
Этот тип резистентности — «жесткий» — более
жест-ко связан со спиралью ареактивности. Такая резистентность является
основой поддержа-ния гомеостаза в пределах уровня граничных параметров
жизни, основой стойких «доно-зологических и нозологических
состояний». Состояние защитных систем и энергетики при этом
напряженное и чревато срывом. Аналогичный тип резистентности часто
встре-чается в старости, в связи с переходом на высокие этажи.
Таким образом, роль состояний ареактивности заключается также в
сохранении гомеостаза, особенно в случаях, когда необходима й возможна
минимальная трата энер-гии: при длительной повышенной активации высоких
уровней реактивности (стойкое фи-зиологическое состояние) и длительном
систематическом стрессе и переактивации, на-против, низких уровней
реактивности (стойкие патологические СОСТОЯНИЯ). В первом случае это
связано с хорошей синхронизацией колебательных процессов, что приводит
к увеличению амплитуды колебаний и частым резонансным перескокам со
спирали реакций на спираль ареактивности и наоборот. Это направлено к
оптимизации состояния. Во вто-ром случае увеличение амплитуды
осуществляется не за счет истинной синхронизации, а за счет
гиперсинхронизации на фоне частичной десинхронизации процессов. Это
жизнен-но необходимо, так как чрезмерные энергозатраты при реакции
соответствующего уровня реактивности могут быстро привести к гибели.
98
Исходя из количественно-качественной закономерности развития системы
адапта-ционных реакций и ареактивности, можно полагать, что у людей,
живущих в естествен-ных (природных) или
антропогенных (возникающих в процессе производственной
дея-тельности ' человека) экстремальных зонах (Авцын А. П., 1966),
развиваются либо неспе-цифические адаптационные реакции на низких
уровнях реактивности (особенно внача-ле/например при переезде на
Крайний Север и т. п.), либо возникают соответствующие
этим реакциям (т. е. также на низких уровнях реактивности —
высоких этажах) состояния ареактивности. Последнее характерно для
живущих уже некоторое время в соответст-вующей «экстремальной
зоне», так как уровень реактивности задается постоянно
дейст-вующим количеством раздражителя, который становится как бы
«фактором среды». Переменные же факторы (или изменение
среды обитания), меняющиеся как по количеству, так и по качеству, до
тех пор, пока они сохраняют свою новизну для организма, вызывают ту или
иную реакцию. Когда «новизна» утрачивается и эти факторы
перестанут вызывать реакцию, от их интенсивности будет зависеть не
характер реакции, а фон существования организма —
уровень реактивности или, вернее,
некоторый набор
уровней.
Так, исследования, проводимые у
жителей Крайнего Севера, выявляют
при-знаки напряжения в
нейро-эндокринной системе («синдром полярного
напряжения», Казначеев В. П., 1980), психоэмоционального
напряжения, сопровождающегося повыше-нием содержания кортикостероидов в
крови (Короленко Ц. П., Бочкаре-ва Н. Л.,
Соко-лов В. П., 1979), изменения в лейкоцитарной формуле и снижение
неспецифической рези-стентности (Сапов И. А., Новиков В. С,
1984), напряжение в сердечно-сосудистой систе-ме, проявляющееся в
развитии гипертонии (Турчинский В. И.), переход с углеводного на
липидный источник энергии (Панин Л. Е., 1983), повышение
уровня заболеваемости
респираторными инфекциями вплоть до эпидемических вспышек
(Деряпа Н. Р., Рябинин И. Ф., 1977) и т. п. Чаще всего эти нарушения
физиологического статуса организма, на-блюдающиеся при переезде в.
«экстремальную зону», авторы рассматривают как проявле-ние
напряжения адаптационной деятельности с переходом в последующем на
новый уро-вень функционирования (Панин Л. Е., 1983; и др.), который,
однако, не является стабиль-ным, так как через некоторое время
наблюдается срыв с ухудшением состояния (Сапов И. А., Новиков В. С,
1984). Описываемое этими и другими исследователями состояние
на-пряжения не укладывается в картину классического стресса
(Данишевский Г. М., 1968; Сороко С. И., 1984), так как отсутствуют
типичные признаки стресса, например, харак-терные для стресса
изменения функции гипоталамо-гипофизарной системы (Василевский Н. Н.,
Сороко С. И., Богословский М. М., 1978).
99
Если проанализировать приведенные данные литературы с позиции
теории адап-тационных реакций и
системы ареактивности, можно увидеть, что речь идет, как уже
говорилось, о «высокоэтажных» реакциях и соответствующих им
уровнях ареактивности, которые отличаются по комплексу изменений от
стресса. Наблюдаются характерные для, этих реакций напряжение
регуляторных механизмов, наличие признаков напряженности в
морфологическом составе крови, метаболизме, снижение резистентности.
Такие реакции чреваты срывом в стресс (или переактивацию) низких
уровней реактивности с развитием различных заболеваний.
Однако имеются данные, которые с позиции роли частотных характеристик
адап-тационных реакций говорят о том, что при хорошей адаптивности даже
в экстремальных условиях уровень реактивности-организма может
повышаться. Так, С. И. Сороко (1984) отмечает, что в Антарктиде у лиц с
плохой адаптивностью происходит резкий сдвиг час-тотного спектра ЭЭГ в
сторону более высоких частот, а при хорошей адаптивности отме-чается,
напротив, усиление медленной электрической активности.
На основании всего сказанного можно предложить профилактику и
оздоровление населения «экстремальных зон» с помощью
целенаправленного вызова и поддержания физиологических адаптационных
реакций (с соответствующим биохимическим сопрово-ждением), т. е.
реакций тренировки и активации особенно повышенных, вызываемых ма-лыми
по абсолютной величине воздействиями. Длительное поддержание реакции
актива-ции может привести к развитию энергетически выгодной
повышенно-активационной аре-активности — стойкого состояния
здоровья, благоприятного также и в плане создания вы-сокой
работоспособности.
Таким образом, периодическая зависимость адаптационных реакций и
состояний ареактивности от величины изменяющегося или постоянно
действующего фактора позво-ляет подойти к управлению функциональным
состоянием организма и его подсистем. Связь периодической системы
реакций и ареактивности с колебательными процессами открывает новые
подходы к такому управлению.
100
ПРЕДСТАВЛЕНИЕ О НОРМЕ, ПРЕДБОЛЕЗНИ, БОЛЕЗНИ, РЕЗЕРВАХ И СТАРЕНИИ ОРГАНИЗМА В СВЯЗИ С РАЗЛИЧ-НЫМИ ТИПАМИ АДАПТАЦИОННЫХ РЕАКЦИЙ
Тип адаптационных реакций и норма
Существует много различных представлений о том, что считать нормой. Мы
под-ходим к представлениям о норме с позиции ее связи с характером
неспецифических адап-тационных реакций организма.
Ранее, говоря об изменениях, характерных для развития различных
адаптационных реакций — тренировки, активации и стресса, мы
сравнивали эти изменения с уровнем ин-тактных животных, которые носили
условное название «контроль». Мы как бы считали контроль
нормой.
Что же такое норма? Мы говорим: реакция тренировки — это реакция
на слабые раздражители (на каждом уровне реактивности), реакция
активации — реакция на раз-дражители средней силы (промежуточные
между слабыми и сильными). Иными словами, и реакция тренировки, и
реакция активации — это реакции, которые встречаются в жизни
постоянно, но не являются чрезвычайными, повреждающими,
патогенными.
Острый стресс, возникающий в ответ на разовое, хотя и чрезвычайное
воздействие, также отнюдь не всегда ведет к развитию болезни и
возникает, когда болезни еще; нет. Поэтому некоторые параметры стресса
также включаются в состав «нормы». Более редко входит в
понятие так называемой нормы хронический стресс, вызванный повторным
дей-ствием чрезвычайных раздражителей. Однако поскольку хронический
стресс может про-текать вяло, не всегда сопровождается выраженным
патологическим процессом, его параметры также частично входят как
составляющая при определении понятия
«норма».
Таким образом, в настоящее время понятие «норма» является
очень сборным и, фактически, включает в себя различные характеристики
стадий реакций тренировки и ак-тивации и даже нерезко выраженного
стресса.
Очень часто исследователи отмечают большую вариабельность нормы,
наличие разнонаправленных изменений внутри нее, большую неоднородность
нормы. Действи-тельно, хорошо известно, что норма — это не
точка, не линия, а всегда — зона, и зачас-тую довольно ш и
р о к а я.
После того, как мы очертили границы изменений, характерные для каждой
адапта-ционной реакции, и научились подбирать дозу, чтобы вызывать
определенную реакцию, мы убедились в том, что
101
разброс в «опытной» группе значительно меньше, чем в
контрольной, в которой животные не подвергались дозированным
воздействиям. По нашему мнению, в понятие
«норма» должны входить только изменения, характерные
для ненапряженных реакций тренировки
и активации.
Тяжелый стресс, по существующим представлениям, является
неспецифической основой болезни, более мягкий стресс —
пред-болезни. Состоянию болезни соответствует также длительная
переактивация. Состоянию предболезни часто соответствуют реакции
тренировки и активации с выраженными признаками напряженности. Это
показано с по-мощью многолетних исследований характера адаптационных
реакций у людей, находя-щихся в различных состояниях (Гаркави Л.
X., Квакина Е. Б., 1987).
Тип адаптационных реакций и резервы организма
Очень часто высказывается мысль, что организм в течение жизни не использует всех своих резервов.
Что такое резервы живого организма?
В самом простом понимании — это тот запас веществ, который может
быть при необходимости переведен в энергию, например, запасы гликогена
в печени, жира — в жи-ровых депо и т. п. Это так называемые
наличные резервы. Эти резервы не являются какой-то неизменной
величиной: постоянно происходит как их использование, так и накопление.
В процессе эволюции, которая привела к созданию высокоорганизованных
организмов со сложной системой регуляции, сформировался другой тип
резервов — функциональные резервы, своего рода запас возможностей
функционирования различных органов и систем. Функциональные резервы
могут быть представлены и в виде определенного структурного запаса
(кровяные депо, избыток капилляров, не включившихся в общий кровоток,
времен-но «молчащие» нейроны в ЦНС), в виде дублирования
физиологических механизмов и, наконец, способности многих органов к
увеличению массы за счет изменения количества функционирующих клеток
при определенных функциональных состояниях. Согласно Г. Н. Кассилю
(1978), потенциальные резервы живой системы в большей степени
определяются возможностью увеличения скорости кругооборота биологически
активных ве-ществ.
Каково состояние резервов организма при разных адаптационных реакциях?
Хо-рошо известно, что при стрессе в стадии тревоги происходит быстрая
мобилизация налич-ных резервов организма. Функциональные резервы
используются только в том направле-нии, которое позволяет быстро
Мобилизовать наличные резервы. Этому содействует и быстрый выброс
адреналина, и повышенная секреция АКТГ и глюкокортикоидов, причем
102
энергия образуется не только наиболее эффективным путем —
окислительного фосфори-лирования, но и путем гликолиза и даже путем
распада белков, РНК и липолиза. Такая не-экономная трата резервов, тем
не менее, целесообразна при ответе организма на сильный раздражитель,
так как помогает
выжить.
В стадии резистентности, благодаря развитию запредельного торможения,
такого быстрого расходования наличных резервов не происходит, напротив,
может происходить даже частичное восстановление запасов —
гликогена, АТФ и др. Однако если действие стрессора продолжается или
стрессор был очень сильным, наступает истощение. Стадия истощения может
длиться более короткое или более длительное время, но так или иначе в
этой стадии распад преобладает над синтезом, происходит постепенное
использование наличных резервов и истощение функциональных, приводящее
к
гибели.
Можно ли заключить, что при этом исчерпываются все функциональные
резервы?
Если не изменить состояние организма, именно так и происходит. Однако
если пе-ревести организм из стадии истощения стресса в реакцию
тренировки или активации, функциональные, а затем и наличные резервы
вновь появляются.
Даже, если судить по распространенной стандартной пробе с АКТГ, в
стадии ис-тощения стресса она свидетельствует об истощении наличных и
потенциальных функцио-нальных резервов коры надпочечников, а после
перевода стресса в реакцию тренировки или активации — о
восстановлении и наличных, и потенциальных резервов. Значит, эти
резервы были не исчерпаны, а скрыты — не могли выявиться без
перевода организма в другое функциональное состояние. Перевод стресса в
реакции тренировки и активации был получен нами специально, с помощью
малых и средних доз нейро-тропных веществ, биостимуляторов и магнитного
поля.
В процессе развития стресса после стадии тревоги наступает стадия
резистентно-сти. Как уже говорилось, она развивается благодаря
преобладанию в мозгу запредельного торможения, снижающего
чувствительность. Если действие стрессора прекращается и стрессор был
не чрезмерно сильным, это запредельное торможение помогает организму
выйти из стресса даже без специальных воздействий, в стадия истощения
не
развивается.
Если действие стрессора было разовым, другие раздражители на фоне
развития запре-дельного торможения будут восприниматься как слабые или
средние, вследствии чего ра-зовьется реакция тренировки или активации.
Выход из стресса с помощью реакции тренировки биологически целесообразен, так как энерготраты при этой реакции малы и посте-
103
пенно происходит накопление энергодающих субстратов — наличных
резервов — самым экономным путем. А это особенно важно после
стресса, сопровождающегося бурной тра-той энергии и наличных резервов.
Если же действие стрессора продолжается, то благодаря сниженной
чувствительности оно может также восприниматься не как сильное
раздраже-ние, а как среднее или даже слабое, также вызывая
соответствующие реакции — актива-ции или тренировки. Именно с
этим мы связываем ту неоднородность и неопределен-ность, которая
имеется в описании стадии резистентности стресса, в отличие от четкости
в описании стадии тревоги и истощения. Можно сказать, что нормализация
наблюдаемых при реакции тревоги сдвигов, иногда даже повышение
некоторых ранее сниженных функ-ция происходит фактически не за счет
стресса, а за счет других приспособительных реак-ций. Если действие
стрессора продолжается, в мозгу исчезает состояние запредельного
торможения и развивается стадия истощения, которая по своим
характеристикам, как уже говорилось, напоминает стадию тревоги.
Фактически без специальных воздействий стресс может перейти в другие
реакции (в которых функциональные резервы используются более
согласованно и эффективно), благодаря развитию запредельного
торможения. Запредельное торможение развивается не только в стадии
тревоги, но и периодически в стадии истощения, а за счет сниженной
чув-ствительности может также происходить переход в другую реакцию без
специального вмешательства. Однако в стадии истощения это явление
кратковременно, если стрессор продолжает
действовать.
Таким образом, становится ясно, что специально вызывать стресс для
лучшего ис-пользования резервов как наличных, так и потенциальных явно
нецелесообразно. К сожа-лению, стресс развивается гораздо чаще, чем это
нужно, и чтобы лучше использовать функциональные резервы организма,
необходимо в первую очередь научиться выводить организм из стресса.
Как обстоит дело с использованием наличных резервов при реакции
тренировки? В отношении использования наличных резервов реакция
тренировки — самая экономная. Энергетические траты малы, уровень
обмена снижен, процессы синтеза я процессы распа-да уменьшены. В
течение стадии перестройки энергетические траты остаются
незначи-тельными, т. е. наличные резервы даже в какой-то степени
увеличиваются. В стадии тре-нированности обмен веществ и энергетические
траты — на самом низком уровне. Иными словами, если говорить о
наличных резервах, они при реакции тренировки не тратятся, а даже
накапливаются. Что же касается функциональных резервов, роль которых
особенно велика, то при развитии реакции тренировки им не грозит
истощение. Это связано с тем, что, во-первых, функциональная активность
регулирующих систем при
104
реакции тренировки умеренна, а во-вторых, с тем, что деятельность
различных систем сбалансирована, нет одностороннего, однобокого
усиления деятельности одной системы при ослаблении других. Поэтому
длительное поддерживание реакции тренировки не при-водит к
истощению резервов.
При реакции активации состояние резервов отличается и от стресса, и от
реакции тренировки. В отношении наличных резервов необходимо отметить,
что происходят вол-нообразные изменения — то трата, то
пополнение. Однако истощения этих резервов не наблюдается. По-видимому,
они пополняются благодаря хорошему состоянию функцио-нальных резервов.
Энергетические траты больше, чем при реакции тренировки, но не так
велики, как при стрессе, так как хорошо сбалансированы с
восстановительными процес-сами. Что касается пластического обмена,
наблюдаются волнообразные изменения. Коле-бания происходят в пределах
верхней половины зоны нормы. Поэтому можно считать и спокойную, и
повышенную активацию анаболическими реакциями, с преобладанием
син-теза. Регуляторные системы — нервная система, эндокринные
железы работают активно и гармонично, нет резкого преобладания
деятельности одних желез над
другими.
Длительное поддерживание реакции активации в наших исследованиях
никогда не приводило к истощению наличных или функциональных
резервов.
Ганс Селье, открывший и исследовавший стресс,
считает, что существует опре-деленное,
отмеренное количество адаптационной энергии,
израсходовав, которую, ор-ганизм больше не может приспосабливаться и
погибает. Если считать, что стресс — это единственная
приспособительная реакция, такое
представление об отмеренном от рождения запасе
адаптационных возможностей можно понять. Действительно, стресс приводит
к более или менее быстрому истощению резервов в зависимости от силы
стрес-сора и длительности его действия, а полноценного пополнения,
резервов при стрессе не происходит. Однако благодаря наличию других
общих неспецифических адаптационных реакций —
физиологических реакций тренировки и активации, а также
благодаря воз-можности перевода организма из стресса
в другую реакцию, «адаптационная
энер-гия», т. е. функциональные резервы организма, могут
пополняться и, следовательно, ока-зываются
значительно большими, нежели можно
предположить. Таким образом, ис-тощенные
резервы могут вновь восстанавливаться, скрытые
резервы становятся явными, и организм вновь и вновь пополняет
свои функциональные резервы. В
настоящее время еще не известно, как велики на самом деле
резервные возможности
организма.
105
Тип адаптационных реакций и старение
Как связан процесс старения с приспособительными реакциями целого организма?
Существует целый ряд теорий старения на клеточном и молекулярном
уровне. Общее в большинстве этих теорий — признание роли в
старении накопления «ошибок» — мутаций в генетическом
аппарате клетки. Однако большинство исследователей признает, что
ста-рение на клеточном и молекулярном уровне происходит медленнее, чем
на уровне целого организма. По словам А. Комфорта (1967), «хвост
кенгуру стареет медленнее, чем сам кенгуру». Почему целый
организм стареет быстрее? Может быть потому, что теряет спо-собность
адаптироваться, приспосабливаться к бесконечным изменениям внутренней и
внешней среды? По мнению В. В. Фролькиса (1969) и его учеников,
организм в процессе старения не теряет способности к приспособлению, но
вырабатывает иные, особые пути приспособления. Фролькису
(1969—1976) и его сотрудникам удалось получить многие интересные
факты о работе различных органов и систем организма, в первую очередь
ЦНС, при старении. Особенно важным нам представляется убедительно
показанный факт повышения при старении чувствительности нервных центров
и, в том числе, центров ги-поталамуса, к возбуждающим сигналам.
Адаптационные реакции, как известно, формируются в ЦНС именно под
влиянием различных гуморальных раздражителей, биологически активных
веществ, образующихся в тех или иных количествах при самых различных
воздействиях на организм. Обычно ин-тенсивность воздействий, падающих
на организм, в течение жизни существенно не изме-няется; не возрастает,
но и не снижается. Исходя из этого, можно предположить, что обычные,
привычные для данного организма раздражения, вызывающие развитие
реакций тренировки или активации—физиологических адаптационных
реакций, при старении мо-гут, становясь относительно более сильными,
вызывать развитие реакции стресса. Дейст-вительно, анализ данных
литературы показывает, что многие изменения при старении на-поминают
изменения, наблюдающиеся при стрессе (Дильман В. М. 1972—1987).
Дильма-ном была предложена так называемая элевационная теория старения,
основанная на том, что активность регуляторных систем, в частности
активность гипоталамического отдела мозга, ведающего регуляцией
внутренней среды организма, с возрастом не снижается, а напротив,
повышается. Это выражается прежде всего в повышении порогов к
гомеостати-ческому торможению. О повышении порогов чувствительности
гипоталамуса к тормозящим воздействиям Дильман судит по возрастному
нарастанию секреции гипофизом го-надотропных гормонов, ослаблению
торможения глюкозой секреции соматотропного гормона, уменьшению от-
106
вета на дексаметазон (не столь значительное, как в норме, уменьшение
экскреции 17-оксикортикостероидов в так называемом «длинном
дексаметазоновом тесте») и т. п.— на-рушениям, в результате
которых чрезмерно усиливается деятельности периферических эндокринных
желез. Факты, приводимые Дильманом, очень убедительны, они
свидетель-ствуют не о нормальном повышении функций гипоталамуса, а о
повышении сверх нормы, характерном, главным образом, для стресса.
Недаром сам Дильман говорит о большом сходстве процесса старения с
различными видами стресса (в том числе и опухолевого), при которых
происходит не только повышение сверх нормы, напряжение деятельности
гипоталамической области мозга, но и нарушение регулирующей
деятельности гипотала-муса, дисбаланс.
Метаболизм изменяется при старении и стрессе также аналогичным образом.
На-блюдаются инсулинорезистентность, снижение толерантности к глюкозе,
накопление жи-ра, усиление липолиза и соответственное повышение уровня
жирных кислот, гипертриг-лицеридемия, гиперхолестеринемия. Эти
нарушения обмена вызывают метаболическую иммунодепрессию (Дильцан В.
М., 1976).
Все это говорит о том, что большую роль в процессе старения на уровне
организма играет преобладание в пожилом возрасте хронического стресса.
Если это так, то показан-ная нами возможность преодоления хронического
стресса открывает пути борьбы с преж-девременным старением на уровне
организма.
Отечественные геронтологи занимают активную позицию в наступлении на
ста-рость. Дильманом (1976) на основании выдвинутой им теории старения
предлагаются оп-ределенные активные вмешательства, нормализующие те
нарушения, которые развивают-ся в регуляционных системах организма при
старении, например, применение антидиабе-тических бигуанидов. Широкую
известность получили рекомендации, связанные с актив-ным отдыхом и
оптимальной физической нагрузкой, как средствами, повышающими
неспецифическую резистентность организма, увеличивающими
продолжительность жизни и нормализующими деятельность основных систем
организма (Аршавский И. А., 1968, 1976; Муравов И. В., 1976;
Маньковский Н. Б. и др., 1971; Чеботарев Д. Ф., Коркушко О. В.,
Калиновская Е. Г., 1969; и др.).
Проведенные в Институте геронтологии АМН СССР комплексные многолетние
ис-следования по изучению действия ряда биологически активных веществ
подтвердили их эффективность в борьбе со старением (Западнюк В. И.,
1984).
Предложены также средства, нормализующие нарушения, развивающиеся при
ста-рении на молекулярном уровне: антиоксиданты (Эмануэль Н. М. 1972),
хелатные агенты (Дубина Т. Л., 1971).
Мы подходим к проблеме борьбы со старением со своих позиций —
107
с позиций необходимости преодоления хронического стресса путем развития
реакции ак-тивации, преобладающей в молодом возрасте (Гаркави Л. X.,
1969; Гаркави Л. X., Квакина Е. В., Уколова М. А., 1974, 1976).
Нами проводились опыты по борьбе со старением на белых крысах-самках в
воз-расте 18—26 месяцев (130 животных) и самцах в возрасте
26—32 месяцев» (50 живот-ных). Реакцию активации вызывали с
помощью электрического раздражения гипоталаму-са, введением 2—20
мкг/кг веса адреналина (подкожно), мумие per os, воздействием
пере-менных магнитных полей индукцией 3,2—4,5 мТ, частотой 50 Гц
на голову животных. С помощью всех этих воздействий у животных
удавалось в большинстве случаев (55—57%) вызывать и поддерживать
реакцию активации в течение длительного времени (2—3 меся-ца).
Через 1-2 недели после начала воздействий появлялись первые видимые
признаки омоложения. По истечении 2—3 месяцев воздействия этих
животных по внешнему виду можно было отличить от молодых только по
размерам. Животные становились более жи-выми, подвижными: редкая,
желтая, грубая шерсть, обычно неопрятного вида, сменялась белой,
мягкой, густой; желтоватые склеры глаз становились ярко-розовыми; кожа
из гру-бой и толстой превращалась в мягкую, эластичную, что напоминает
изменения, разви-вающиеся у животных с калорийно недостаточным питанием
(Никитин Н. В., 1976). Из-менения в организме были характерными для
реакции активации: тимус увеличен, соот-ношение нейтрофилов и
лимфоцитов — как при спокойной или повышенной активации,
функциональная активность щитовидной железы умеренно повышена, секреция
минера-локортикоидных гормонов коры надпочечников преобладала над
секрецией глюкокорти-коидных без снижения последней. Мы полагаем, что
комплекс изменений; в эндокринной системе, описанный у животных,
длительно сохранивших молодость в результате приме-нения сдерживающего
рост питания (Никитин В. Н., 1976; Мороз Ю. А., Никитин В. Н.,
Ставицкая Л. И., 1976) и названный авторами «мягким
стрессом», является фактически не стрессом, а именно активацией.
Длительное применение активации приводило к нормализации нарушенного
поло-вого цикла. Если к моменту начала воздействия у самок 18—22
месяцев в части случаев преобладал эструс, в части диэструс, а у самок
свыше 26 месяцев — анэструс, то через 3—5
недель систематического воздействия начиналось восстановление
нормального поло-вого цикла (рис. 42). Необходимо подчеркнуть, что
такой эффект наблюдался в тех случа-ях, когда удавалось получать и
стойко поддерживать в организме реакцию активации. По-сле прекращения
воздействий уже через 3 месяца у части животных, а через 6 месяцев
— у всех нарушения полового цикла возобновлялись,
и начинал изменяться их внешний
вид. Через полгода
108
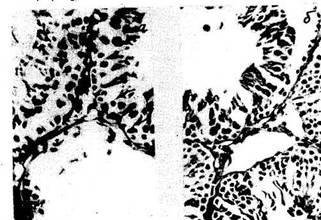
Рис. 42. Изменение полового цикла у старых крыс-самок при развитии стойкой реакции активации.
а - контроль (1), нарушение цикла (2); б - динамика полового цикла: 1,
3- нарушенного (при стрессе) и 2, 4- восстановленного (при активации)
109
у 29 животных трехмесячный курс активации был повторен с таким же
результатом. Это говорит о том, что активационные курсы
«лечения» должны систематически
повторяться.
У самцов также через 2—3 недели после начала лечения отмечалось
повышение половой активности. Морфологические исследования, проведенные
у части животных, показали, что в семенниках наблюдалась характерная
для реакции активации стимуляция сперматогенеза. После прекращения
воздействий, так же, как у самок, уже через 3—6 ме-сяцев
происходили обратные изменения. Проведенный у 21 самца повторный курс
дал тот же положительный эффект (Гаркави Л. X., 1969; Гаркави Л. X.,
Квакина Е. Б., Уколо-
ва М. А., 1979).
Таким образом, опыты на животных говорят о возможности
целенаправленного лечения старения.
Необходимо подчеркнуть, что эффективность
лечения зависела не столько от вида применяемых воздействий, сколько от
того, как удавалось поддерживать реакцию
активации.
На основании полученных нами данных мы полагаем, что в плане
оптимизации тех воздействий, которые применяются для активного
вмешательства в процессы старения, очень важно контролировать характер
развивающейся адаптационной реакции. Хорошо известно, например, что в
то время, как умеренные физические упражнения приводят к обратному
развитию признаков преждевременного старения, к расширению диапазона
ре-акций организма (Муравов И. В., 1976), большие физические
нагрузки оказывают небла-гоприятное влияние и в эксперименте приводят
даже к уменьшению продолжительности жизни животных. В то же время
многочисленные данные говорят о том, что ограничение двигательной
активности, гипокинезия вызывает преждевременное старение. В. В.
Фроль-кисом (1975)1 отмечена наибольшая продолжительность жизни
подопытных животных при умеренных раздражениях. Как увеличение, так и
уменьшение силы раздражений снижало
продолжительность жизни. По нашему мнению, благоприятное действие
уме-ренных раздражении может быть связано с развитием реакции
активации.
1
Имеющиеся клинические наблюдения о влиянии биологически активных
препара-тов говорят о том, что эффект действия этих веществ в
зависимости от дозы различен (Свечникова Н. В., Беккер В. И., Мороз Е.
В., 1976). Мы считаем, что оптимизация дозы биологически активных
веществ под контролем реакции активация улучшит эффект этих
препаратов.
Таким образом, представления о норме, резервах организма старении во
многом связаны с типом адаптационных реакций.
110
ПРЕДСТАВЛЕНИЕ О НОРМЕ, ПРЕДБОЛЕЗНИ, БОЛЕЗНИ, РЕЗЕРВАХ И СТАРЕНИИ
ОРГАНИЗМА В СВЯЗИ С МНОГОУРОВНЕВОСТЬЮ РАЗВИ-ТИЯ АДАПТАЦИОННЫХ РЕАКЦИЙ И
АРЕАКТИВНОСТИ
Что изменилось в представлениях о норме, предболезни и болезни после
того, как выяснилось, что по мере изменения силы (дозы)
воздействия адаптационные реакции тре-нировки, спокойной активации,
повышенной активации и стресса закономерно цикличе-ски, периодически
повторяются на разных уровнях реактивности?
Норма, предболезнь и болезнь с учетом реакций
разных уровней реактивности и состояний ареактивности
Как уже говорилось, уровень реактивности определяется абсолютной
величиной действующего фактора, (мг, мл, мТ и т. п.). Если действующий
фактор большой по абсо-лютной величине, реакция развивается на низких
уровнях реактивности (высокие этажи). Если действующий фактор небольшой
по абсолютной величине, реакция развивается на высоких уровнях
реактивности (низкие этажи). На каждом уровне реактивности слабое для
данного уровня воздействие вызывает развитие реакции тренировки,
среднее — раз-витие реакции спокойной или повышенной активации,
сильное — стресса. Иными слова-ми, упоминаемые ранее понятия
«слабое», «среднее» и «сильное»
являются в какой-то ме-ре относительными, так как относятся к данному
уровню реактивности. Поэтому' реакцию тренировки (реакцию на слабый
раздражитель) высокого этажа можно вызвать дейст-вующим фактором,
большим по абсолютной величине, чем стресс (реакцию на сильный
раздражитель) низкого этажа. Необходимо подчеркнуть, что периодическая
система адап-тационных реакций имеет четкую количественную
характеристику — коэффициент реак-ции.
Таким образом, вместо четырех реакций (тренировки, спокойной активации,
по-вышенной активации и стресса) имеется многоуровневая система этих
реакций (Гаркави Л. Х., Квакина Е. Б., 1975— 1987).
Если проанализировать комплекс изменений в организме при развитии
адаптаци-онных реакций тренировки, активации и стресса на разных
уровнях реактивности, будет ясно, что представлениям о норме,
здоровье соответствуют реакции активации и трени-ровки высоких уровней
реактивности. Именно при этих реакциях отмечается высокая
функциональная активность защитно-приспособительных механизмов,
гармоничность, т. е. хорошая синхронизация деятельности различных
систем, отсутствие напряженности и признаков полома,
повреждения.
111
Действительно, хорошая адаптация, высокая
резистентность организма, отсутствие элементов повреждения, гармоничная
работа различных систем организма, умеренные энергетические траты
— норма, здоровье; плохая адаптация, низкая резистентность,
нали-чие элементов повреждения, дисгармония в работе различных систем
организма, большие и неэкономные энерготраты — предпатология,
предболезнь, патология, болезнь.
Понятие нормы должно как можно больше соответствовать понятию здоровья.
Ведь нормативы и определяются для того, чтобы малейшие отклонения от
них служили сигналом тревоги: нужно вмешаться, чтобы вернуть здоровье.
С этой точки зрения наше представление о норме согласуется с
представлениями В. М. Дильмана об «идеальной норме»—
уровне гомеостаза, свойственного 20—25-летнему возрасту, когда в
подавляю-щем большинстве случаев встречается реакция активации (чаще
— повышенной) высоких уровней реактивности.
Исходя из этих представлений, можно сказать, что условиям,
предъявляемым к по-нятию «норма», соответствуют только
реакции активации и тренировки высоких уровней реактивности, т. е.
вызываемые малыми по абсолютной величине воздействиями. При этом
нет элементов напряженности, отдельные системы организма работают
согласован-но, гармонично. Реакция активации высоких уровней
реактивности часто встречается, как уже говорилось, в молодом здоровом
организме.
Что касается болезни, то со времени первых работ Г. Селье, которому
принадлежит одно из ярких определений стресса как «симптома
болезни вообще», стресс несет на себе бремя ответственности за
все болезни. Учитывая многоуровневую систему адаптацион-ных реакций,
можно частично реабилитировать стресс, так как стресс высоких уровней
реактивности не является столь грубо повреждающим. С другой стороны,
появилось представление о переактивации — состоянии, во многом
противоположном стрессу (дру-гая крайность). Итак, на данном уровне
знаний, в рамках предложенной классификации, можно заключить, что
состояние болезни может быть описано в параметрах стресса и
пе-реактивации, развивающихся на низких уровнях реактивности.
Что же осталось на долю предболезни? Реакции тренировки и активации
низких уровней реактивности и стресс — высоких. Эти реакции, не
имеющие еще черт явного повреждения, болезни, но приобретающие элементы
напряженности и рассогласования работы подсистем организма, уже
содержат в себе угрозу здоровью.
Предложенные простые критерии верификации адаптационных реакций по
лейко-цитарной формуле были использованы для оценке уровня здоровья в
различных коллек-тивах: при массовых профилактических осмотрах, среди
занимающихся закаливающими
112
процедурами и физической культурой, в группах людей, предъявляющих
различные жа-лобы, и т.
п.
Обследование молодых здоровых людей, здоровых людей среднего возраста,
детей и части здоровых людей пожилого возраста показало, что в
большинстве случаев у них регистрируется реакция активации повышенная
(чаще) или спокойная и реже — реакция тренировки (Бутов М. А.,
1984; Сауткин М. Ф., Поляков А. П., Иванова М. В., 1984; Глух-ман М.
В., Ступин Г. К., 1979; Ногаллер А. М., Бутов М. А., Калыгина Т. А.,
1984), Эле-менты напряженности в лейкограмме при этом отсутствовали,
что позволило отнести эти реакции к физиологическим, низкоэтажным.
Иными словами, реакции тренировки и акти-вации без элементов
напряженности, развивающиеся на высоких уровнях реактивности организма,
являются неспецифической основой нормы, здоровья. При этих реакциях,
осо-бенно при реакций активации (в первую очередь, повышенной),
деятельность желез внут-ренней секреции гармонична и соответствует
норме молодых и здоровых, работают сис-темы, ответственные за иммунную
защиту, отмечается высокая работоспособность, со-стояние центральной
нервной системы — без явлений перераздражения или угнетения.
Именно при этих «низкоэтажных» реакциях, особенно при
повышенной активации — ус-тойчивом состоянии здоровья —
человек наиболее способен справляться со стрессом.
Обследование людей, предъявляющих различные жалобы на состояние
физическо-го и психического дискомфорта, утомляемость, снижение
работоспособности, иногда го-ловные боли и боли в области сердца,
подверженность сезонным простудным заболевани-ям, плохой сон и аппетит
и т. п., хотя и не имеющих как будто определенного заболева-ния,
показало, что у них чаще всего встречаются либо напряженные,
дисгармоничные ре-акции тренировки и активации, т. е. развивающиеся на
высоких этажах, либо мягкий «низкоэтажный» стресс — с
незначительной лимфопенией и без резких признаков напря-женности. Лишь
у небольшой части (около 5%) параметры лейкограммы укладывались в рамки
гармоничных, физиологических реакций. Скорее всего, в этом случае
реакции раз-вивались также на высоких этажах, хотя и не имели
выраженных признаков напряженности по лейкоцитарной формуле.
В целях массового обследования населения предлагаются различные
критерии оценки здоровья и распознавания донозологических состояний,
хотя, как правило, они ха-рактеризуют состояние не организма в целом, а
какой-либо из его подсистем — сердечно-сосудистой, дыхательной и
т. д.
Включение метода верификации неспецифических адаптационных реакций в
ком-плекс мероприятий по обследованию рабочих и служащих промышленных
предприятий, наряду с методами оценки состояния сердечно-сосудистой
системы и высшей нервной
113
деятельности, показало, что гармоничные реакции тренировки, спокойной и
повышенной активации распределялись следующим образом: у 19% отмечалась
реакция тренировки, у 24,1%—спокойная активация, у 26,2%—
повышенная активация; у 8,6% отмечался хрони-ческий стресс, у
21,2%— напряженные реакции тренировки и активации. Как отмечают
авторы, особый интерес представляло сопоставление результатов скрининга
по состоянию сердечно-сосудистой си- схемы с результатами по
исследованию адаптационных реакций: было обнаружено совпадение групп
«напряженные» по тому и другому критериям (Фи-липченко Р.
Е., Привалов И. А., Любарская К. И., Алексеева Г. П., 1980).
Таким образом, простые критерии адаптационных реакций могут быть
применены при массовых осмотрах, в целях диспансеризации, для выявления
состояния предболезни.
Что привносит понятие «состояние ареактивности» в представление о норме и па-тологии?
Как уже было показано, состояниям ареактивности свойственна стойкость и
все стойкие состояния связаны с ареактивностью. Пока известно три
стойких состояния: одно из них — стойкое состояние здоровья, оно
связано с повышенноактивационной ареактив-ностью низких этажей и внешне
проявляется как стойкая повышенная активация. Рези-стентность при этом
состоянии очень высока, и eго нужно отнести к одному из лучших
вариантов нормы. Два других стойких состояния связаны со стрессорной и
переактиваци-онной ареактивностью высоких этажей и проявляются при
обычных исследованиях как довольно глубокий стресс и резкая
переактивация. Эти состояния являются неспецифиче-ской основой
различных заболеваний.
Таким образом, рассмотрение представлений о норме, предболезни и
болезни с по-зиций многоуровневой системы адаптационных реакций и
ареактивности вносит новое в понимание этих категорий в теоретическом
плане. Выяснилось, что понятиям не только болезни, но и предболезни и
даже здоровья соответствует не одно, а несколько; различных
состояний, связанных с развитием различных
адаптационных реакций на разных уров-нях реактивности, а также с
состояниями ареактивности. От того, какие это реакции и уровни
реактивности и связаны ли они с ареактивностью, зависит большая или
меньшая стойкость как состояний здоровья, так и болезни. Значительное и
стойкое отклонение го-меостаза характеризует состояние,
являющееся неспецифической основой
болезни, что полностью совпадает с
представлениями В. М. Дильмана
(1987)
Поскольку имеются простые показатели реакций и, в какой-то степени,
уровни ре-активности по компонентам лейкоцитарной формулы, как
основному — процентному со-держанию лимфоцитов, так
и сопутствующим
(другие элементы
лейкоцитарной формулы
114
а также число лейкоцитов), можно составить представление, к какой
категории относится человек в данное время. О стойкости состояния можно
судить, повторяя эти простые ис-следования в динамике. Следовательно,
этот подход может быть полезен в практике как для диагностики
состояний, так и для их коррекции в процессе лечения.
Резервы организма с учетом реакций
разных уровней реактивности и ареактивности
Если ранее, когда было известно о существовании адаптационных реакций
на од-ном уровне реактивности, мы говорили, что стресс способствует
лишь трате резервов, а реакция тренировки и активации открывают пути к
их восстановлению, то наличие реак-ций на разных уровнях реактивности
позволяет подойти к этому вопросу более диффе-ренцированно. Хотя стресс
разных уровней реактивности действительно приводит к пре-обладанию
расходования резервов — и наличных, и функциональных, и
энергетических, — но величина расходования очень различна.
При стрессе Г. Селье — стрессе высоких этажей (низких уровней
реактивности), когда на организм действует большой по абсо-лютной
величине раздражитель, идет быстрое и значительное расходование
резервов. При мягком стрессе — более высоких уровней реактивности
(низких этажей), при действии раздражителей не таких больших по
абсолютной величине (являющихся лишь относи-тельно сильными для своих
уровней реактивности) . также преобладает расход резервов, но степень
его существенно , меньше. Так, например, если при тяжелом стрессе масса
ти-муса 20—30 мг (а иногда тимус даже трудно найти) на 100 г веса
крысы, то при более мягком стрессе масса тимуса может быть 70—80
мг (на 100
г).
При реакции тренировки — той реакции, которая в энергетическом
отношении яв-ляется наиболее экономичной, способствующей , накоплению
энергетических резервов (например, фонд эндогенной янтарной
кислоты), а процессы анаболизма и катаболизма пластических веществ
находятся на низком уровне, происходит восстановление резервов.
Однако выраженность восстановления и накопления резервов различная и
находится в за-висимости от уровня реактивности
(этажа): на низких этажах (большая
абсолютная вели- чина действующего фактора) эта способность
уменьшается. Например, масса тимуса при реакции тренировки высокого
этажа 80—90 мг на 100 г веса, а при реакции трениров-ки низкого
этажа — до 170— 180 мг.
При реакции активации как спокойной, так и повышенной, когда расход и
накоп-ление энергии хорошо уравновешены и наблюдается даже накопление
пластических ве-ществ (особенно при повышенной активации),
восстановление резервов происходит осо-бенно быстро.
115
Однако и в этом случае многое зависит от уровня реактивности:
накопление и восстанов-ление резервов происходит лучше также на высоких
уровнях реактивности (низких эта-жах), т. е. при реагировании на малые
по абсолютной величине действующие факторы. Например, масса тимуса при
развитии реакции активации на высоких этажах достигает 180—190 мг
при спокойной и 200—220 мг (на 100 г веса крысы) при повышенной;
на низ-ких этажах масса тимуса— 280—300 мг при спокойной и
350—370 мг при повышенной
активации.
Приводя в виде примера массу тимуса, мы хотим подчеркнуть, что самые
главные резервы — это не наличные, а функциональные, 3
функциональным же изменениям всегда соответствуют определенные
структурные (Саркисов Д. С. 1987).
Изучение морфологических изменений многих органов и тканей при реакциях
раз-личных уровнях реактивности показало, что по мере
возрастания абсолютной величины
действующего фактора число структур, участвующих в
реакции, возрастает. Это совпа-дает с со временными представлениями
(Саркисов Д. С, 1987; Меерсон Ф. 3., 1986; и др.) о структурной цене
адаптации. Однако поскольку при различных реакциях (тренировки,
спокойной активации, повышенной активации и
стрессе) изменения различны,
это увеличение числа структур носит нелинейный характер. Мы
согласны с тем, что увеличе-ние участвующих в реакции структур можно
характеризовать как «мобилизацию налич-ных материальных ресурсов
органа» (Д. С. Саркисов), но, как оказалось, такая мобилиза-ция
чаще находится не в прямой, а в
обратно пропорциональной зависимости от уровня
резистентности. По-видимому, это связано с большими тратами
энергии, а уро-вень резистентности очень
четко зависит от энергетической цены. Поэтому
восстанав-ливать и накапливать резервы, как это ни парадоксально
на первый взгляд, лучше с по-мощью воздействий, малых по абсолютной
величине, которые энергетически экономич-ны, легче выводят из стресса и
вызывают развитие реакции активации.
Старение с учетом реакций разных уровней
реактивности и
ареактивности
Как уже говорилось, мы связывали процесс старения с более частым
развитием стресса в пожилом возрасте, когда обычные раздражители
становятся как бы более силь-ными. Известно, что стресс ускоряет
процесс старения (Selye H., Tuchweber В.,
1976).
Дальнейшие исследования у лиц разного возраста, в том числе! пожилых и
старых, пока-зали, что в большом возрасте встречаются не только стресс,
HOW реакции тренировки, спокойной и повышенной активации.
Вместе с тем в лейкограмме пожилых и старых
116
людей очень часто имеются элементы напряженности. Кроме того, изменение
соотноше-ния глюкокортикоидных гормонов коры надпочечников и гормонов
щитовидной железы у пожилых характерно для реакций, развивающихся на
высоких этажах (низких уровнях ре-активности), т. е. при действии
больших по абсолютной величине раздражителей. Поэто-му в настоящее
время можно представить себе, что по мере старения привычные для
каж-дого организма раздражители (даже без увеличения внешних нагрузок),
становясь относительно более сильными, сдвигают реакции организма на
все более и более высокие этажи («вверх по лестнице, ведущей
вниз»), на которых реакции, как отмечалось ранее, являют-ся
энергетически менее выгодными, дисгармоничными, напряженными. В этом же
на-правлении, еще более закономерно, согласно В. М. Дильману, действуют
и онтогенетиче-ские механизмы — онтогенетический путь старения по
Дильману. Согласно этим пред-ставлениям, старение является побочным
результатом онтогенетической программы развития, закона отклонения
гомеостаза.
Развитие адаптационных реакций на высоких этажах имеет своим следствием
об-наруженное Дильманом при старении и других «нормальных
болезнях» повышение гипо-таламического порога к торможению.
Дильман пишет: «Старея, человек и вне стресса на-чинает жить, как
бы находясь в состоянии хронического стресса, и поэтому становится все
более и более беззащитным, когда реальный стресс предъявляет свои
требования к орга-низму» (Четыре модели медицины. М., 1987. С.
98). Этот «как бы стресс» является по сути и стрессом, и
другими реакциями, но развивающимися на высоких этажах (низких уров-нях
реактивности), на которых эти реакции, будучи напряженными, легко
переходят в стресс.
Таким образом, полностью разделяя выдвинутую Дильманом концепцию о
законе отклонения гомеостаза, мы рассматриваем ее с наших позиций как
закон постепенного повышения этажа (снижение уровня реактивности), т.
е. выбор организмом в процессе старения все больших по абсолютной
величине раздражителей. Переход на реакции высо-ких этажей —
реакции с элементами напряженности, дисгармонии, большими
энергетиче-скими тратами создает закономерное отклонение гомеостаза в
процессе старения.
В реализации и закона отклонения гомеостаза В. М. Дильмана, и
закона сохране-ния гомеостаза К.
Бернара существенную роль играет периодическая система
адапта-ционных реакций и ареактивности. Оба эти закона составляют
диалектическое единство, соответствующее принципу устойчивого
неравновесия Э. Бауэра.
Закономерность пе-рехода на реакции низких уровней реактивности
и одновременно реализация закона
отклонения гомеостаза могут быть объяснены с позиции
представлений об организме как о сложной
колебательной системе. Любые
колебания на разных
117
иерархических уровнях представляют собой отклонения от точки, условно
называемой точкой покоя. По мере роста и развития организма происходит
увеличение массы тела, секреции различных гормонов и т. п., т. е.
осуществляется интенсивные рост и развитие с использованием
положительной обратной связи. С позиции представлений о роли
в жиз-недеятельности организма колебательных
процессов это говорит об увеличении откло-нения от точки покоя,
что неизбежно приводит к
изменению положения самой точки
покоя — сдвигу ее в сторону максимума колебаний, т. е. отклонению
от прежнего уровня (в данном случае — уровня гомеостаза).
Это особенно наглядно на примере изу-чения энергетического обмена (на
уровне митохондрий) — при резком повышении скоро-сти дыхания
происходит невозвращение к исходной точке покоя, тем более
выраженное, чем было большим первоначальное
отклонение. Это характерно, в первую очередь, для стресса,
предъявляющего к организму большие требования, но в какой-то степени
прису-ще и другим реакциям, развивающимся на низких уровнях
реактивности. Когда програм-ма роста заканчивается и организм
достигает «идеальной нормы» по Дильману, этот
про-цесс не прекращается, тем более, что он поддерживается и
наличием больших внешних возмущений.
Если исходить из этих представлений, для борьбы со старением нужно
вызывать не просто реакцию активации, а реакцию активации высоких
уровней реактивности, т. е. с помощью очень малых
раздражителей. Это тем
более целесообразно, что
при ста-рении уменьшаются различия в чувствительности между
женскими и мужскими особями: и те, и другие становятся более
чувствительными, т. е. способны реагировать на целена-правленно
применяемые малые по величине раздражители. Кроме того, уже давно было
отмечено, что биологическое время нерегулярно, и, например, при
старении, болезни и тяжелых нагрузках оно течет
быстрее. Мы связываем «ускорение»
биологического вре-мени с высокой частотой колебаний при развитии
реакций на низких уровнях реактивно-сти (высоких этажах) и
большими энергетическими тратами. И напротив, в молодом
здо-ровом организме реакции (особенно
реакция активации) развиваются на энергетически выгодных высоких
уровнях реактивности с меньшей частотой колебаний при выраженном
негэнтропийном эффекте и неравновесности, что может способствовать
замедлении; тече-ния времени (Гаркави Л. X., Квакина Е. В., 1984;
Гаркави Л.X Квакина Е. В., Лупичев Л. Н., 1985). Это также говорит в
пользу роли минимизации воздействий при лечении старе-ния.
Было проведено 5 серий опытов на 316 старых крысах самцах и самках
весом более 300 г в возрасте 24—26 месяцев и самцах в возрасте
26—32 месяцев с воздействиями ад-реналином в дозе 0,2 мкг/кг
веса, норадреналином 0,5—0,2 мкг/кг и переменным магнит-ным полем
индукцией 0,1—0,7 мТ частотой 50 Гц. Дозу (силу) воздействия
118
варьировали около указанных в пределах
±20% с уметом адаптационных
реакций (коррекция по лейкоцитарной формуле). У 92%
всех старых животных исходно отмечал-ся стресс, а у 8% —
адаптационные реакции тренировки и активации с элементами
напряженности: эозинопения, лейкоцитоз,
лейкопения. Регулярное исследование ваги-нальных мазков у
старых крыс-самок показало, что к моменту начала воздействия у самок в
64% наблюдался диэструс, а у остальных — эструс; другие стадии
подового цикла отсутствовали, т. е. отмечались
четкие нарушения полового цикла. С помощью перечис-ленных
слабых воздействий у 78% подопытных животных удалось вызывать и стойко
поддерживать адаптационные реакции активации и тренировки (активацию в
65% случа-ев). Полное восстановление полового цикла у самок при
развитии реакции активации и омоложение
животных по внешнему виду (отличие от молодых
только по размерам) происходило не через 2—3 месяца, а уже через
5 недель. В тех случаях, когда преобла-дающей была реакция
тренировки, также отмечалось восстановление нормальной
дли-тельности полового цикла
и последовательности его стадий,
однако продолжи-тельность отдельных стадий не нормализовалась. Так,
наблюдалась большая длительность метэструса, меньшая длительность
эструса и проэструса. В тех случаях, когда развитие реакций активации и
тренировки получить не удавалось, и преобладающей реакцией был стресс,
восстановления общей продолжительности полового цикла
не отмечалось, по-следовательность стадий
цикла продолжала оставаться нарушенной. Однако, в отличие от
контрольных животных без воздействий, у подопытных животных происходило
вос-становление стадии проэструс, хотя длительность ее была
меньше, чем в норме. По-видимому, разница со стрессом в контроле
связана с тем, что под влиянием воздействий, малых по абсолютной
величине, развивался стресс низкого этажа.
При гистологическом исследовании яичников старых крыс, переведенных в
реак-цию активации различными слабыми воздействиями, в корковой
части было выявлено небольшое количество фолликулов на разных
стадиях созревания (от примордиальных до зрелых). Отмечались единичные
желтые тела малых размеров с хорошо сохранившейся структурой
лютеоцитов. Наблюдалась значительная пролиферация клеток интерстиция,
среди которых выявлены очаговые скопления тучных клеток, богатых
Шик-позитивным альцианофильным метахроматическим материалом. Так же,
как и в яичниках, в маточных трубах
периваскулярная соединительная
ткань характеризовалась усилением
Шик-позитивного окрашивания, альцианофилии и метахромазии с Азуром-1.
Гистологическая картина яичников старых
крыс контрольной группы была иной. Обращало внимание наличие
большого количества желтых тел крупного размера. Грани-цы лютеоцитов
определялись
119
нечетко, и ядра находились в состоянии пикноза. В узких прослой
ках интерстиция и стенках сосудов
отмечалась очаговая эозино
филин..
Анализ морфологического исследования семенников у старых крыс-самцов
после систе-матического поддержания реакции активации с помощью малых
доз адреналина свиде-тельствовал о выражен ном адаптационно-трофическом
влиянии на процессы сперматогенеза. Это проявлялось в увеличении числа
фигур митоза в пресперматидах, усилении ок-раски ядер сперматоцитов,
повышении количества сперматозоидов в апикальных частях клеток Сертоли,
а также вакуолизации интерстициальной ткани. В то же время при
разви-тии стресса наблюдалось значительное увеличение числа
дегенеративно измененных из-витых канальцев, накопление в них детрита,
атрофия стенок канальцев, что свидетельст-вует об угнетении функции
семенников (рис. 43) (Гаркави Л. X., Квакина Е. Б., Уколова М. А., 1979;
Рис. 43. Семенник старой крысы: а — реакция стресс: атрофия части
канальцев, снижение спермато-генеза; б — реакция активации:
восстановление нормальных структур на фоне еще оставшейся час-тичной
атрофии стенок канальцев, усиление сперматогенеза, вакуолизация
интерстициальной ткани. Видны фигуры митоза. Окр. гематоксилин-эозином.
Ув. 40X7
Шихлярова А. И., 1983; Шихлярова А. И., Мулатова А. К., Евстра-това О. Ф., Мамцева Л. И., 1984).
Кроме того, отмечалось увеличение тимуса со стимуляцией лимфоидной
ткани, по-вышение функциональной активности щитовидной железы и
минералокортикоидной функции коры надпочечников. Признаки омоложения
после прекращения малых по абсо-лютной величине воздействий держались,
дольше (чем при действий
120
больших) на 3—4 месяца. Активационные курсы лечения, повторенные
у части животных (126), дали не менее выраженный, чем первый курс,
положительный эффект. У небольшой части животных, которых было возможно
оставить для наблюдения, длительность жизни (в плохих условиях вивария)
колебалась от 4 (3 животных) до 5 лет (4 животных).
Полученные результаты говорят о большей эффективности малых по
абсолютной величине воздействий, которые не только вызывают развитие и
стойкое поддержание ре-акции активации — реакции, наиболее
способствующей омоложению, но и переводят ор-ганизм на высокие уровни
реактивности, где реакция активации является болееe гармо-ничной и
экономичной.
Общеизвестно, что отклонения гомеостаза при
старении у разных индивидуумов выражены в различной степени, причем
разница эта может быть весьма существенной. Чем она объясняется? На
основании проведенных многолетних исследований, можно ска-зать, что это
связано и с этажом, и с реакцией. Что значит связано с этажом? Это
значит, с той абсолютной величиной раздражителя, которую
преимущественно выбирает организм для реагирования. Как уже говорилось
ранее, молодой здоровый организм может выбирать малые раздражители из
множества падающих на него. Способность выбирать ма-лые действующие
факторы присуща организму в реакции активации, особенно повышен-ной,
высоких уровней реактивности. Переходя в процессе жизни на высокие
этажи, орга-низм теряет способность выбирать малые по абсолютной
величине раздражители, однако способность реагировать на малые
раздражители полностью не утеряна, и применяя их целенаправленно, можно
навязать организму реакций высоких уровней реактивности, о чем
свидетельствуют опыты по лечению старения. Как уже говорилось, выбор
раздражи-теля определенной величины связан с резонансными явлениями и
разной частотой коле-баний, присущих реакциям на разных уровнях
реактивности.
Исследование ферментов лимфоцитов (дегидрогеназ) молодых животных (10
меся-цев — 1 год), стареющих (1,5-2 года) и старых (3—3,5
года) без учета характера адаптаци-онных реакций показало, что эти
изменения носят фазный характер. У стареющих по сравнению с молодыми
отмечалось значительное повышение активности всех изучавших-ся
ферментов (в 1,5-2 раза) и увеличение числа жестких корреляционных
связей между ними, что свидетельствует о большом напряжении
энергетических процессов. В группе старых животных, напротив,
наблюдается существенное снижение активности всех фер-ментов, особенно
СДГ, не только по сравнению со стареющими, но и с молодыми живот-ными.
Одновременно отмечается ослабление корреляционных связей между ними.
Когда же сравнительный анализ активности ферментов был проведен с уче-
121
том характера адаптационных реакции. оказалось, что эти фазные
изменения особенно выражены у животных в стрессе, в то время как у
животных, находящихся в активации, отсутствуют достоверные возрастные
изменения активности всех ферментов, кроме не-большого уменьшения
активности СДГ в группе старых животных (Гаркави Л. X., Кваки-на Е. Б.,
Котляревская Е. С. Марьяновская Г. Я., Мамцева Л. И., 1981). Нужно
подчерк-нуть, что эти исследования проводились у животных без
специального вызова реакции активации, т. е. активация была спонтанной
и, следовательно, довольно высокоэтажной (не специально вызванной
малыми раздражителями). Таким образом, по этим параметрам у
«активационных» животных не отмечалось существенных
отклонений от уровня гомео-стаза молодых
животных.
Исследования содержания в крови холестерина, сахара, гормонов
щитовидной же-лезы, артериального давления в пожилом и старом
возрасте также показали, что у лю-дей с
преобладающей реакцией активации эти изменения укладываются в
картину, близ-кую к «идеальной
норме». Большой интерес
представляет исследование тех гомео-статических
тестов, которые предлагаются Дильманом (1987) для паспорта
здоровья. Однако но всем уже проведенным исследованиям —
функционального состояния
ЦНС, эндокринных желез,
тимико-лимфатической системы,
некоторых показате-лей метаболизма
— ясно, что реакция активации, особенно повышенная, главным
обра-зом, высоких уровней реактивности, является
своеобразной реакцией антистарения, то-гда как
стресс, напротив, реакцией старения.
Учитывая важность воспроизведения потомства для сохранения жизни вида,
не вы-зывает сомнений мысль о существовании специальной программы
развития, направлен-ной на создание оптимальных условий для размножения
(Дильман В. М., 1987). Не вызы-вает также возражений, что закон
отклонения гомеостаза является следствием программы развития, и
старение (как и сопутствующие ему «нормальные» болезни)
является побоч-ным результатом этой же программы. Однако, естественно
предположить наличие других программ сохранения жизни, не связанных с
размножением, особенно по отношению к высшим организмам, индивидуальный
опыт которых приобретает важное значение для развития и сохранения
вида. Вообще, сохранение жизни — более общая цель, а размно-жение
— лишь один, хотя и важный, даже доминирующий на данном отрезке
эволюции живого, путь сохранения жизни. С этой точки зрения нам
представляется, что антистрес-сорная система неспецифических
адаптационных реакций запрограммирована в процессе эволюции именно для
сохранения жизни вообще, а не только для создания оптимальных условий
для размножения. В такой сложной, многомерной и вместе с тем надежной
сис-теме, как живой организм, мало-
122
вероятно наличие лишь одной программы сохранения жизни. Обычно любой
вид деятель-ности организма обеспечивается многими путями работой, как
минимум, двух систем (система и антисистема по Г. И. Крыжановскому,
1987), а чаще — многими системами. Это видно на примере
многоуровневой системы реакций, где антистрес сорную систему составляют
и реакция повышенной активации с выраженными антистрессорными
харак-теристиками, и реакция спокойной активации, где антистрессорные
признаки выражены менее резко, и реакция тренировки, которая имеет и
черты сходства, и черты различия со стрессом. По-видимому, программы,
направленные на сохранение жизни, как индивиду-альной, так и жизни
вида, также неоднозначны и многообразны. Возможность спонтанно-го
развития реакции активации при старении в настоящее время снижена,
по-видимому, вследствие гиподинамии в сочетании с большими
психоэмоциональными стрессорными нагрузками, избыточным и
нерациональным питанием. Однако целевая программная функции реакции
активации как механизма замедления старения и, вообще, отклонений
гомеостаза очевидна. Целенаправленный вызов и поддержание в организме
реакции акти-вации в те возрастные периоды. когда В. М. Дильманом
отмечается нарушение идеальной нормы (т. е. после 20—25 лет),
может явиться реальным способом задержки старения и связанных с ним
«нормальных» болезней. Это тем более реально, что реакция
активации имеет последействие, так что проводиться курсами
активационной профилактики. Кроме того, уже разработаны
программированные режимы (с учетом возраста и пола) вызова ре-акции
активации на высоких уровнях реактивности (без необходимости
систематического исследования даже такого простого показателя, как
лейкоцитарная формула крови). По нашему мнению использование в
программированных режимах препаратов, предлагаемых Дильманом для борьбы
с отклонением гомеостаза, будет особенно полезно, так как позво-лит
сочетать специфический и неспецифический механизмы антистарения.
Немаловажное значение для реализации программы сохранения жизни имеет и
система ареактивности, хотя этот вопрос еще мало изучен и нуждается в
дальнейших ис-следованиях. Однако уже сейчас можно сказать, что на
поздних этапах старения ареак-тивность довольно высоких этажей
становится важным путем повышения резистентное™ организма. Этот
путь не самый эффективный и экономичный, но он препятствует откло-нению
гомеостаза, которое приводит организм к большому напряжению. Этим путем
дос-тигается продление жизни в старости. По-видимому, с этим
связана фаза глубокого ста-рения, когда многие показатели
жизнедеятельности (например, содержание различных гормонов), до этого
повышенные напротив, снижаются. Наиболее стойким состоянием
здоровья и одновременно
антистарения является
повышенно-активационная
123
ареактивность высоких уровней реактивности.
Это состояние является наиболее экономичным по энерготратам и наиболее
негэн-тропийным из всех известных нам в настоящее время состояний
организма. Если удается его поддерживать у пожилых и старых людей, то
это способствует оздоровлению, хоро-шему самочувствию, замедлению
старения и сохранению работоспособности даже в глу-бокой
старости.
1
ПРИНЦИПЫ
ПРИМЕНЕНИЯ
I
РЕАКЦИЙ АКТИВАЦИИ И ТРЕНИРОВКИ ДЛЯ ПРОФИЛАКТИКИ И ЛЕЧЕНИЯ
Как было показано, вызывая
какую-либо адаптационную реакцию, мы факти-чески вызываем в
организме определенный комплекс изменений в деятельности основных его
подсистем, создавая определенное функциональное состояние целого
организма. В зависимости от того, какая реакция, т. е. какой комплекс
изменений развивается, сопро-тивляемость организма
повышается или понижается. Если целенаправленно
вызывать реакцию, повышающую резистентность, устойчивость
организма к повреждающим, факторам, таким
образом можно и оздоравливать организм,
и проводить профилак-тику, и лечить, причем лечить
разные болезни. Это, конечно, не панацея — болезни
раз-ные, и таким путем
этиологического лечения осуществить нельзя. Но,
вместе с тем, все болезни реализуются в организме, и
от его состояния, активности его защитных под-систем зависят
уровень здоровья, устойчивость к
болезням и нагрузкам, тяжесть за-болевания и
даже эффективность специфического лечения.
Является ли лечение с помощью повышения устойчивости организма чем-то прин-ципиально новым? Безусловно, нет.
Еще во времена Гиппократа было замечено, что, с одной стороны,
большинство заболева-ний может излечиваться без посторонней помощи, а с
другой — выздоровление невоз-можно без усилий самого организма.
Эти представления легли в основу учения о целеб-ных силах организма.
Можно сказать, что принципы неспецифической терапии были за-ложены еще
в VI в. до н. э. Одним из основных правил Гиппократа было лечение в
соот-ветствии с индивидуальными особенностями. Этот принцип был так
сформулирован Ари-стотелем: «Если кто владеет понятием, а опыта
не имеет и общее познает, а заключенного в нем индивидуального не
ведает, такой человек часто ошибается в лечении, ибо лечить приходится
индивидуальное». Кроме того, Гиппократ подчеркивал значение
мероприя-тий, воздействующих на весь организм (например, диета, напитки
и вообще образ жизни), так как «человек един». Согласно
Гиппократу, лечение не только должно быть индивиду-альным, но и должно
меняться в течение заболевания у одного и того же больного.
124
Наиболее выдающиеся врачи последующих веков следовали принципам лечения
Гиппократа. К ним относятся крупнейшие отечественные клиницисты: М. Я.
Мудров, Г. А., Захарьин. С. II. Боткин. «Употребляя различные
методы лечебные, не думайте только, что вы лечите самую суть болезни.
Врач лишь помогает организму справиться с болезнью, справиться его
собственными защитными приемами. Все это можно осуществить исходя из
требования - брать организм в целом и одновременно учитывая влияние
окружающей среды на организм человека»,— писал Боткин. Н.
В. Лазаревым было показано, что изме-няя общий уровень деятельности
защитных систем организма, влияя фармакологическим путем на
«предсуществующие в организме комплексные защитные реакции и
механиз-мы», можно вызвать в организме состояние неспецифически
повышенной сопротивляемо-сти (CHПС). Лазарев подчеркивал большую
перспективность такого направления но срав-нению с попытками влиять на
отдельные звенья в цепи нарушений, развивающихся в ор-ганизме,
например, при таком тяжелом заболевании, как опухолевая болезнь.
Но почему этот способ лечения не стал преобладающим? Но может быть
лучше, чем лечение силами самого организма! В настоящее время известно,
каково значение со-противляемости организма при самых различных
заболеваниях, в том числе инфекцион-ных и даже опухолевых болезней. В
то же время, применяемая до на стоящего времени неспецифическая терапия
витаминами, биостимуляторами различного происхождения, средствами
иммунотерапии в одних случаях высокоэффективна, в других — не
оказывает существенного влияния, а иногда оказывает даже
нежелательное влияние.
По нашему мнению, основной причиной неудач различных видов
неспецифической терапии
является чисто качественный
подход к действию различных веществ, без специального учета
количественной стороны, без учета того, что одно и то же средство
в различных дозах может оказать на организм разное даже прямо
противоположное дейст-вие. В медицине имеется пристрастие к
жестким схемам лечения, установлению одинако-вых дозировок без учета
индивидуальной чувствительности
организма. Для дозиров-ки
учитываются в основном только физические
параметры организма: вес, поверх-ность тела, объем
циркулирующей крови и т. д.
По нашим представлениям, благодаря разной индивидуальной
чувствительности одинаковые дозы вызывают у одних развитие ре акции
тренировки, с помощью которой за короткий срок не удается значительно
повысить резистентность организма и которая ока-зывает мягкое
противовоспалительное действие, у других — реакцию активации,
быстро и значительно повышающую резистентность и одно-
125
временно провоспалительный
потенциал, а у третьих —
развитие стресса, что может привести даже к ухудшению
состояния.
Успех и неуспех
неспецифической терапии, проводимой
без количественно-го подхода, является в значительной мере случайным
Даже при иммунотерапии, обосно-ванной теоретически, при подборе
дозы учитывается лишь
специфическое действие на
исследуемые факторы иммунитета. Однако развитие специфических и
неспецифиче-ских защитных реакций
тесно связано между
собой. Мы полностью согласны с Н. В.
Васильевым (1973, 1975), что разделение
раздражителей на специфические и
неспецифические (в иммунологическим смысле)
является условным, так как, с одной стороны, в ответ на действие любого
антигенного раздражителя в организме развивается
комплекс неспецифических изменений,
а с другой - различные
раздражители неан-тигенной природы
(вибрация, звук магнитное поле) вызывают в лимфоидной ткани
изменения, напоминающие иммуноморфологический комплекс.
«Нам не удалось найти таких клеточных звеньев
лимфоидной ткани, которые бы реагировали только на
анти-ген и не отвечали на неантигенное воздействие» (Васильев Н.
В., 1975). Согласно Василь-еву, неспецифическая клеточная реакция в
ответ на введение антигена является фунда-ментом, основой формирования
специфического иммунитета. Имеются также данные,
свидетельствующие о глубокой внутренней
связи адапто- и иммуногенеза (Васильев Н. В., Коляда Т. И.,
1987).
Как уже говорилось, мы также получили при развитии реакции активации (с
помо-щью адреналина и магнитного поля) изменения, характерные для
антипенного воздейст-вия (Гаркави Л. X., Кваки-на Е. В., Уколова М. А.,
Мулатова Л. К., Шихлярова А. И., 1982; Мулатова А. К., 1983). На
основании этого мы думаем, что и при иммунотерапии дозу воздействия
целесообразно подбирать по показателям той адаптационной реакции,
которую необходимо вызвать в данном случае. В пользу такой
необходимости свидетель-ствуют разнородные результаты, получаемые при
иммунотерапии.
Для того, чтобы вызывать развитие адаптационных реакций, нужно хорошо
пони-мать, что почти каждое воздействие в зависимости от его силы,
биологической активности может вызывать различные неспецифические
адаптационные реакции. Конечно, специфическое повреждающее действие
отдельных раздражителей делает их непригодными для использования с этой
целью. Так, с помощью разового облучения животных дозой 0.2-0,5 Гр
удалось вызвать развитие реакции активации с характерным для нее
комплексом изме-нений (Гаркави Л. X., Мацанов А. К., 1971). Однако вряд
ли целесообразно вызывать ре-акцию
активации облучением, особенно повторным.
Не отрицая значения качества воздействия,
мы хотим еще раз подчеркнуть, что одно и то же но
качеству воздействие может вы-
126
звать разные изменения в организме. Мы подчеркиваем это потому,
что в медицинских работах часто злоупотребляют чисто качественным
подходом к оценке
воздействий: адреналин вызывает и
гипергликемию, и повышение артериального давления, магнитное поле
гипокоагуляцию (и кровотечение)
и т. д. Между тем адреналин,
в дозах, вы-зывающих реакцию активации, приводит
даже к гипогликемии и умеренному сниже-нию артериального
давления, магнитное поле вызывает гипокоагуляцию лишь при разви-тии
реакции тренировки и РПА и т. д. С другой стороны, в оценке
результатов во многих медицинских работах преобладает
количественный подход: больше — меньше. Это ве-дет к
неправильной трактовке результатов, особенно если исследуется один
показатель, а не соотношение между различными (пусть
и связанными друг с другом)
показате-лями. Так, например, и в первую фазу стресса, и в первую фазу
реакции тренировки про-исходит
повышение секреции глюкокортикоидов, хотя
и различное по степени. Сек-реция же
минералокортикоидов впервой фазе стресса угнетена, а в первой фазе
реакции тренировки умеренно увеличена. Разное
соотношение этих гормонов обусловливает четкую
разницу между реакциями. Об этом же говорит разная степень
повышения секре-ции глюкокортикоидов:
количество пере ходит в качество. Если же судить только по
принципу «больше-меньше», не
учитывая количественно-качественных
переходов и соотношений в изменении
различных показателей, то совершенно разные
состояния могут показаться одинаковыми. И напротив, одни и те же
состояния могут трактоваться как различные. Так, в процессе смены
стадий реакций тренировки и активации происхо-дят фазные
изменения различных показателей —
то повышение, то снижение.
На-пример, содержание
глюкокортикоидов, повышенное в
первую фазу реакции тре-нировки, в
третью уже снижается до исходного уровня.
Использование количествен-но-качественного принципа приспособления,
который развился в процессе эволюции, по-зволит наиболее целесообразно
применить действие самых различных факторов для по-вышения
резистентности организма.
Проведенные исследования показали, что хотя адаптационные реакции можно
вы-звать разными воздействиями, при тяжелых заболеваниях (например,
опухолевой болез-ни) вряд ли кому-нибудь придет в голову использовать
спортивные тренировки, водные процедуры и другие такого рода
общеукрепляющие воздействия, очень полезные для здо-ровых. Наиболее
эффективными из испытанных нами средств для подъема резистентно-сти у
больных были различные нейротропные вещества, биостимуляторы и
магнитное по-ле. Конечно, нами не испытано все бесчисленное множество
возможных воздействий. По-этому не исключено, что в дальнейшем можно
найти и другие достаточно эффективные средства. По нашему мнению, при
выборе средств нужно учитывать, но меньшей мере, два условия.
127
Первое — отсутствие в используемых дозах специфического
повреждающего дей-ствии на отдельные системы организма. Второе
способность оказывать влияние на ЦНС и, особенно, на гипоталамическую
область, в которой происходит формирование адаптаци-онных реакций
тренировки и активации. Благодаря тому, что подкорковые отделы мозга
обладают значительной инерционностью, в случаях длительного поддержания
определен-ной реакции последняя приобретет своеобразную
«инерцию», и другие, случайные, воз-действия (кроме
сверхсильных) не изменяют ее.
Как уже говорилось, в настоящее время хорошо известно, что в основе
многих па-тологических процессов лежит развитие разной степени
выраженности хронического стресса. Поэтому задачей неспецифической
терапии чаще всего является выведение орга-низма из состояния
хронического стресса.
Как можно вывести организм из стресса и перевести в реакцию тренировки
и акти-вации, если стресс — это реакция на сильный раздражитель,
а реакция тренировки и акти-вации — соответственно на слабый и на
средний, иными словами, как более слабые воз-действия могут преодолеть
действие более сильных? На этом вопросе мы останавливались уже подробно
при рассмотрении роли колебательных процессов в развитии всех
этих со-стояний. При тяжелом стрессе в стадии истощения далеко не
всегда удается вызвать и особенно поддерживать реакцию активации, а
реакцию тренировки, которая требует наи-меньших энергетических затрат,
вызвать гораздо легче. В таких случаях иногда приходит-ся поддерживать
некоторое время реакцию тренировки, создавая некий «запас»,
и только после этого удается перейти к реакции активации. Облегчает
перевод стресса в реакцию активации и так называемое
«биохимическое сопровождение» (Кондрашова М. Н., 1975). В
ряде опытов получение противоопухолевого эффекта при больших опухолях
удавалось при сочетании обычных активирующих воздействий с введением
глюкозы и вдыханием большего, чем обычно, количества кислорода.
Особенно эффективной оказалась приме-няемая нами по предложению
Кондрашовой янтарная кислота (Терапевтическое действие янтарной
кислоты. Пущино, 1976).
Вопрос, почему все-таки удается перевести организм в определенную
реакцию с помощью специальных воздействий в то время, как на него
падает множество самых раз-нообразных раздражений, тоже освещался нами
ранее, с позиций роли резонансных явле-ний в развитии адаптационных
реакций. Мы думаем, что случайные слабые и средние раздражители также
выводят организм из стресса, иначе стресс встречался бы еще чаще.
Однако эффект разовых или не систематически повторяющихся воздействий
весьма крат-ковременен. Для достижения стойкого эффекта
необходимо систематическое повторение
воздействий, причем проводимое
под контролем иссле-
128
дований, свидетельствующих о развитии, и
поддерживании в организме той или иной адаптационной
реакции. Контроль необходим потому, что чувствительность
одного ор-ганизма не только отличается от другого (индивидуальная
чувствительность), но у одного и того же организма она волнообразно
меняется с течением времени. Так, если развивает-ся реакция тренировки,
чувствительность меняется волнообразно, но
с направленно-стью к
снижению, и поэтому для под
держания той же реакции силу (дозу)
после-дующих воздействий большей частью нужно немного
увеличить. Если же мы вызываем реакцию активации,
чувствительность, напротив,
несколько повышается (также вол-нообразно), и для
поддержания той же реакции силу последующих воздействий чаще все-го
нужно немного уменьшить. Таким образом, для того,
чтобы вызывать одну и ту же реакцию, необходимо гибко варьировать
силу (дозу) воздействия. Если же доза остается.
одинаковой на протяжении длительного времени, происходит
волнообразная смена адап-тационных реакций
(рис. 44). Мы связываем это с характером изменений в ЦНС,
свой-ственным каждой адаптационной
реакции. При реакции
тренировки в мозгу пре-обладает
Рис. 44. Характер зависимости между величиной применяемой дозы и
развивающимися адапта-ционными реакциями (по числу лимфоцитов): а
- изменение числа лимфоцитов при постоянной дозе; б —
относительное постоянство числа лимфоцитов при
волнообразно меняющейся дозе.
129
состояние охранительного торможения, при реакции активации
— умеренного физиоло-гического возбуждения, а при стрессе —
либо резкого возбуждения, либо запредельного торможения. Поддерживать
длительно одну и ту же реакцию необходимо, чтобы сделать ее стойкой.
Как уже говорилось, адаптационные реакции формируются в подкорковых
от-делах мозга, которые относятся к медленно действующим системам с
большой инерцион-ностью. Систематическое повторение воздействий,
вызывающих определенную реакцию, придает ей инерцию, стойкость.
Вначале надо подобрать дозу (силу) воздействия, необходимую для каждого
инди-видуума, а затем корригировать ее по мере продолжения воздействий.
Подбирать дозу для человека с помощью систематических гормональных и
других сложных исследований практически невозможно. Необходим более
простой, доступный тест для контроля за пра-вильным подбором дозы.
Сопоставление комплекса гормональных исследований с изме-нением числа
лейкоцитов и лейкоцитарной формулы подтвердило уже известные данные о
том, что белая кровь является своеобразным гормональным зеркалом.
Факторами, обес-печивающими возможность ориентировки по белой крови,
является индивидуальная, а не усредненная оценка изменений, строгое
соблюдение необходимой стандартизации усло-вия взятия анализа, и,
наконец, при применении повторных воздействий, возникновение
«инерции» намечающегося сдвига. Последнее приводит к
уменьшению обычно наблю-даемого размаха колебаний в белой крови.
Использование этих тестов на человеке выяви-ло дополнительные
корреляции изменений белой крови с субъективным состоянием
(на-строение, общая активность, сон, аппетит).
Лечение с помощью развития первой фазы реакции тренировки испытано нами
при острых гастритах, колитах, обострении язвенной болезни, бронхитах,
неспецифических инфекционных полиартритах. Это лечение значительно
более мягкое, чем большими, стрессорными дозами глюкокортикоидов
(сопровождающееся иммунодепрессией и сни-жением резистентности) и
достаточно эффективное.
Реакция тренировки может быть использована также для защиты от
повреждающих аген-тов различной природы, особенно тех. действие которых
сопровождается развитием лей-копении. При развитии лейкопении следует
отдать предпочтение реакции тренировки пе-ред реакцией активации. С
помощью первой стадии реакции тренировки было получено мягкое и в то же
время быстрое антикоагуляционное действие. Это особенно эффективно,
когда одновременно необходимо получить и провоспалительное действие
(например, ост-рые тромбофлебиты).
Лечение с помощью развития реакции активации успешно испытано нами при
хро-нических вялотекущих воспалительных процессах (гастриты,
панкреатиты, колиты, яз-венная болезнь желудка
130
и двенадцатиперстной кишки, бронхиты,
неспецифические пневмонии). Реакция
ак-тивации была использована; для
долечивания острых воспалительных
процессов, для защиты
от действия различных повреждающих
факторов при отсутствии
вы-раженной лейкопении (лучевая и химиотерапия опухолей,
применение больших доз про-тивотуберкулезных
препаратов, антибиотиков). Реакция
спокойной активации при-менялась
для нормализации
свертывания крови, а реакция повышенной активации
— для антикоагуляционного действия при хронических тромбофлебитах.
Реакция активации и, особенно, повышенной активации, были применены при
ле-чении злокачественных опухолей. Уже давно замечено, что какое-то
повышение активно-сти защитных систем позволяет организму эффективно
бороться с опухолью.
Реализация противоопухолевого эффекта при реакции активации происходит,
как уже описывалось, путем уничтожения клеток опухоли
лимфоцитами, врастанием
со-единительной ткани в опухоль, т.
е. иными словами, сам
процесс рассасывания опухоли говорит о
повышении иммунной защиты, особенно неспецифического клеточного
иммунитета.
Нами совместно с клиницистами разработан метод лечения
злокачественных опу-холей путем развития и поддерживании реакции
активации с помощью магнитного поля (аппарат «Полюс 1).
Способ заключается в том, что магнитное поле
применяется под контролем систематических анализов
крови (два раза в неделю) и гормональных
исследо-ваний (один раз в месяц). Использовалось
магнитное поле с индукцией от 12 до 45 мТ.
частотой 50 Гц, форма тока
синусоидальная, режим аппарата
прерывистый (авт. свид. № 522688, Панков А. К., Уколова М.
А., Квакина Е. Б., Гаркави Л.Х. Соловьева Г. Р., Еремин В. И.,
Салатов Р. Н., 1976).
Применение этого метода на людях показало, что принципиальной
разницы меж-ду опухолями человека и
животных нет: в тех случаях, когда удается вызывать и
под-держивать реакцию активации происходит регрессия
опухоли. Получена полная
рег-рессия рака нижней губы и кожи I — II стадии. При
этих локализациях в подавляющем большинстве случаев (328 из 360)
удалось получить полное рассасывание
опухоли без применения
других методов лечения. Регрессию
поверхностных опухолей удавалось получить, хотя и более медленно, и при
наличии чередования общего мягкого стресса с реакцией тренировки.
Противоопухолевое влияние при этих опухолях было связано и с местной
реакцией, а именно с развитием местного стресса, имеющего, как и общий
стресс, черты повреждения, дегенерации,
дистрофических процессов, в данном
случае, на уровне клеток опухоли. В то же
время, процесс регрессии опухоли лучше
(быстрее) протекал на
фоне общей реакции
активации, местным проявлением
которой является
лимфоцитарная инфильтрация опухоли.
131
Работы P. П. Кинута с сотрудниками (1986) показали, что магнитные поля
могут быть успешно применены в нейроонкологии в комплексном лечении
злокачественных глиом. Авторы подчеркивают необходимость проверки
индивидуальной чувствительности опухолевой ткани к магнитному нолю (с
помощью авторадиографии или определения ДНК на сцинтилляционном
счетчике).
Вызывать и, особенно,
поддерживать реакцию активации, не
говоря уже о реакции повышенной активации, у больных с опухолями
внутренних органов оказалось значительно труднее. В
зависимости от локализации
опухоли присоединяются нару-шения,
связанные с выключением (частичным или полным) функции
пораженного опу-холью органа. В
каждом конкретном случае необходимо
разрабатывать систему
биохимического сопровождения, чтобы
в какой-то мере преодолеть
нарушения, препятствующие развитию в организме
реакции активации. Метод новый, вполне естест-венно, что он нуждается в
дальнейших исследованиях и разработке. Даже такие давно
применяемые методы лечения опухолей, как лучевая
терапия и химиотерапия, требуют дальнейшего развития
и разработки. Мы считаем очень важным, что принцип
противо-опухолевого действия реакции активации
подтвердился и в клинике. Опухоли
внут-ренних органов, наблюдение за динамикой которых значительно
труднее, чем при наруж-ных опухолях, и опасность метастазирования
которых несравненно больше, мы не имеем права длительно лечить новым
методом без применения старых, проверенных.
Об эффективности активационной терапии при опухолях внутренних органов
сви-детельствует применение активационной магнитотерапии в
предоперационном периоде у больных раком легкого III — IV стадии.
Применяли аппарат «Полюс-1», интенсивность МП варьировали в
пределах 10 — 25 мТ, ежедневную экспозицию — в пределах 3
— 20 мин для получения реакции активации. В процессе
лечения отмечалось улучшение обще-го состояния, а при
рентгенологическом исследовании в 1/3 случаев — уменьшение
опу-холи, почти в половине случаев — стабилизация
(отсутствие роста), у остальных — не-большое увеличение. Однако
последующее (после оперативного вмешательства)
морфо-логическое исследование показало, что увеличение опухоли
произошло за счет массивно-го развития грубой волокнистой
соединительной ткани и
лишь у двух больных
было истинным. У этих больных на протяжении всего
периода лечения был стресс. Морфологическое исследование показало, что
в ткани опухоли определялись разнообраз-ные дистрофические изменения,
вплоть до лизиса опухолевых клеток, существенно более выраженные, чем
встречаются в ткани опухоли без лечения
(Огородникова Л. С, Гай-рабедьянц Н. Г., Ратнер
О. П., Чирвина Е. Д., Гаркави Л. X., Квакина Е.
Б., Уколова М. А., 1980).
Не вызывает сомнений, что хронический стресс, сопровождающийся психиче
132
ской депрессией, в том числе эмоциональной, играет роль в
возникновении, росте и разви-тии онкологических заболеваний (Bahnson С.
В., Bahnson M. В., 1966; Selye H., 1979, 1981; Bаltrusch H. .J. Е.,
Strangel W., 1980; Ваmmег К., 1980; Balitsky К. Р., Vinnitsky V. В.,
1981; Балицкий К. П., Шмалько Ю. П., 1986: Ва-stecky et al., 1984;
Stress and cancer, 1981). Мно-гочисленные уже в настоящее время данные
свидетельствуют о том, что рак можно отне-сти к психосоматическим
заболеваниям (Bastecky et al. 1984. и др.). Изучение механизмов
эмоционального стресса выявило его роль в нарушении процессов
саморегуляции гомео-стаза (Судаков К. В., 1981), в том числе нарушении
иммунного гомеостаза (Крыжанов-ский Г. Н., Магаева С. В., 1983).
Последнее понятно в плане специального участия нерв-ных структур
гипоталамуса и гормонов гипоталамо-гипофизарно-надпочечниковой сис-темы
в регуляции иммунного гомеостаза (Корнева Е. А., Клименко В. М., Шхинек
Э. К., .1978). Известно, что состояние психической депрессии ухудшает
прогноз ракового про-цесса, в то время как при положительном
эмоциональном настрое и активной жизненной позиции отмечается улучшение
прогноза и увеличение длительности жизни даже в далеко зашедших стадиях
рака (Grossath-Maticek R., 1980; Дильман В. М., 1983). Уже эти
обстоя-тельства говорят в пользу необходимости всеми возможными
способами стремиться к борьбе с опухолевым стрессом к целенаправленному
развитию реакции активации, повы-шающей психическую активность и
улучшающей психоэмоциональный статус.
Сравнение трудностей достижения эффекта при лечении с помощью развития
фи-зиологических адаптационных реакций онкологических и
неонкологических заболеваний показало, что при самых различных
заболеваниях эффект получить несравненно более легко, В настоящее время
этот метод (активационной терапии) используется с целью оз-доровления,
профилактики и повышения эффективности лечения различных заболеваний не
только нами, но и его последователями в различных областях медицины. Об
этом будет рассказано специально в следующей главе.
ПРИНЦИПЫ ИСПОЛЬЗОВАНИЯ РЕАКЦИЙ АКТИВАЦИИ И ТРЕНИРОВКИ РАЗНЫХ УРОВНЕЙ РЕАКТИВНОСТИ ДЛЯ ОЗДОРОВЛЕНИЯ, ПРОФИЛАКТИ-КИ И ЛЕЧЕНИЯ
Что меняет в представлении об использовании реакций тренировки и
активации с целью профилактики и лечения знание периодической
закономерности повторения этих реакций на разных уровнях реактивности?
Очень многое. И в эксперименте, и даже в
133
клинике мы получали хорошие результаты потому, что вызывали реакции
тренировки и активации сравнительно небольшими по величине действующими
факторами, т. е. вызы-вали развитие этих реакций на высоких уровнях
реактивности (низких этажах). Как уже говорилось, реакции спокойной,
повышенной активации и реакция тренировки на низких уровнях
реактивности (высоких этажах), когда они вызываются довольно большими
по величине действующими факторами, являются неспецифической основой не
нормы, здо-ровья, а предболезни, иногда даже болезни. На низких уровнях
реактивности эти реакции становятся энергетически менее выгодными,
появляются элементы рассогласования в дея-тельности отдельных
подсистем. Поэтому использование таких реакций для оздоровле-ния,
профилактики и лечения нецелесообразно. Отсюда вытекает очень простой
принцип применения адаптационных реакций с этой целью — принцип
минимизации абсолютной величины действующих факторов. Доза (сила)
должна быть минимальной, но действую-щей.
Исследования показали, что женщины любого возраста сохраняют высокую
чувст-вительность, способность реагировать на малые по абсолютной
величине действующие факторы. Это объясняется, по-видимому, присущим
женщинам циклическим характером изменений в нейроэндокринной системе.
Мужчины же, начиная, примерно с 25-летнего возраста, начинают терять
способность реагировать на малые по абсолютной величине факторы.
Поэтому может возникнуть ситуация, когда воздействие применяется, а
орга-низм на него не реагирует, т. е. создается лишь видимость
воздействия. Как уже говори-лось, организм может выбирать из множества
падающих раздражителей либо малый по величине (при исходных реакциях
высоких уровней реактивности, особенно повышенной активации), либо
большой (при исходных реакциях низких уровней реактивности, осо-бенно
стресса и переактивации) управляющий действующий фактор. Учитывая
важную роль колебательных процессов в формировании основных
функциональных состояний организма, мы связываем такой выбор с
резонансными взаимодействиями организма и действующих на него факторов.
В связи с этим, начиная воздействие, необходимо вы-явить либо по
анализу крови, либо хотя бы по самочувствию, управляется ли организм
применяемым воздействием, т. е. происходит ли развитие нужных реакций
или нет. Целе-направленно переводя организм на реакции высоких уровней
реактивности (низких эта-жей) с помощью постепенного снижения силы
(дозы) действующего фактора, можно вер-нуть ему эту способность. Таким
образом, организм не суммирует все падающие на него воздействия, а
выбирает какое-либо одно, которое тем самым становится управляющим.
Иными словами, организм реагирует дискретно. Для того, чтобы
управляющим стало применяемое воздействие, необходимо соблюдение
некоторых условий. Во-первых, оно
134
должно быть применено в определенное время суток, обычно в промежутке
между 7 и И часами, когда у большинства людей происходит формирование
реакций околосуточного ритма (Баевский Р. М., 1977). При этом, если
воздействие очень мало — фактически, име-ет информационное
значение,— его лучше применять в промежутке между 7 и 9 часами.
Разумеется, это справедливо не для всех. Есть люди с измененными или
даже переверну-тыми ритмами — тут нужно подстраиваться
индивидуально.
Кроме того, учитывая наличие быстрой нервной фазы развития реакции и
более медленной — гуморальной, нами с целью вызова реакции
активации применено двойное воздействие, в котором учитываются
биоритмические особенности обеих фаз.
Важным фактором, способствующим успешному развитию реакций, является
«фактор новизны». Поэтому в одном из способов вызова
реакции активации используется специальный подбор дозы (силы) с учетом
этого фактора.
Наши исследования и данные Ю. В. Серегина (1986) показали наличие
экспонен-циальной зависимости между силой (дозой) воздействия и
характером адаптационной ре-акции. Эта зависимость также была учтена
для создания способа вызова реакции актива-ции.
Наиболее употребительным в течение многих лет был способ вызова и
поддержи-вания нужной адаптационной реакции кибернетически, с
использованием для обратной связи такого сравнительно простого
показателя реакций, как соотношение форменных элементов в специально
подсчитанной формуле.
Разные способы вызова реакции активации применяются для оздоровления,
профи-лактики и лечения (в комплексе со специфическим лечением)
различных заболеваний.
Повышение сопротивляемости организма
с помощью адаптационных реакций тренировки и активации на разных
уровнях ре-активности организма (активационная терапия)* на основе
обратной связи с орга-низмом
Необходимо иметь в виду, что, как уже говорилось, реакции активации и
трениров-ки высоких уровней реактивности без признаков напряжения
являются неспецифической основой нормы; реакции активации (РСА и РПА) и
тренировки низких уровней реактивности и стресс высоких уровней
реактивности — неспецифической основой предболезни; стресс низких
уровней реактивности, переактивация и
реакции активации и
трени-ровки низких уровней
_________________________________
* Выдержки из Методических рекомендаций (составлены Л. X. Гаркави, Е.
Б. Квакиной, М. А. Уколовой, Н. П. Захаровой, Ф. М. Захарюта, А. И.
Шихля ровой), утвержденных МЗ РСФСР 17 января 1978 г.
135
реактивности с резко выраженными элементами напряженности — неспецифической ос-новой патологических процессов.
Наиболее простые показатели адаптационных реакций. Сложные
нейроэндок-ринные изменения, характеризующие каждую из адаптационных
реакций, получают оп-ределенное отражение в морфологическом составе
белой крови. Это дает возможность использовать простые показатели для
каждой реакции, а следовательно, осуществлять контролируемую
неспецифическую активационную терапию. Тип реакции определяется, прежде
всего, по процентному содержанию лимфоцитов в лейкоцитарной формуле.
Ос-тальные форменные элементы белой крови и общее число лейкоцитов,
являясь лишь до-полнительными признаками реакций, свидетельствуют о
степени полноценности реакции, степени ее напряженности и отношении к
общепринятым границам нормы.
Реакция тренировки. Число лимфоцитов в пределах нижней половины зоны
нор-мы (21 — 27%), число сегментоядерных нейтрофилов — в
пределах верхней половины зоны нормы, число палочкоядерных нейтрофилов,
эозинофилов, моноцитов и лейкоци-тов—в пределах нормы.
Реакции активации.
1. Реакция спокойной активации (РСЛ): число лимфоцитов — в
пределах верхней половины зоны нормы (28—33%), сегментоядерных
нейтрофилов — в пределах нижней половины зоны нормы,
палочкоядерных нейтрофилов, эозинофилов, моноцитов — в пре-делах
нормы, лейкоцитов — (4,0—8,0) . 109.
2. Реакция повышенной активации (РПА): число лимфоцитов выше нормы
(более 33, до 40—45% — индивидуально), сегментоядерных
нейтрофилов — ниже нормы, па-лочкоядерных нейтрофилов,
эозинофилов и моноцитов — в пределах нормы, лейкоцитов
--(4,0-8,0) . 109.
Острый стресс. Как известно из работ Г. Селье, острый стресс
характеризуется лейкоцитозом, анэозинофилией, лимфопенией и
нейтрофилезом. Наши наблюдения по-зволили определить примерные
количественные параметры острого стресса у человека: число лимфоцитов
— менее 20%, сегментоядерных нейтрофилов — выше нормы,
палоч-коядерных — норма и выше, эозинофилов — 0, моноцитов
— норма и выше, лейкоцитов — более 8,0 . 109.
Хронический стресс. Число лимфоцитов — менее 20%, сегментоядерных
нейтро-филов — выше нормы, палочкоядерных — норма и выше,
число эозинофилов может быть и малым (до 0), и нормальным, и выше
нормы, моноциты — в пределах нормы, и выше, и ниже, число
лейкоцитов может быть и снижено, и повышено, и в пределах нормы.
Переактивация. Число лимфоцитов — более 40 или 45% (в зависимости
от инди-видуальных особенностей), сегментоядерных нейтрофилов —
ниже нормы, другие фор-менные элементы — в пределах нормы.
136
Основное соотношение для характеристики реакций — между числом
лимфоцитов и сегментоядерных нейтрофилов (ЛФ/СН) — имеет
наибольшую величину при РПА, да-лее в убывающем порядке — РСА,
реакция тренировки, стресс. Нужно иметь в виду, что это соотношение
отражает тип реакции лишь при отсутствии признаков напряженности
реакции. Повышение числа моноцитов для каждой реакции
часто свидетельствует о достижении ее
верхней границы.
Признаки напряженности, неполноценности реакции. Отклонение от
указанных для реакции тренировки и активации параметров белой крови
— лейкоцитоз, лейкопения, эозинопения или эозинофилия, моноцитоз,
монопения, базофилия, сдвиг влево — говорит о неполноценности
данной реакции, о нарушении гармоничности в функционировании подсистем
организма.
Если число моноцитов повышается (более 7%) или снижается (менее 4%),
число эозинофилов либо снижается (менее 1,0%), либо повышается (более
6%), число лейкоци-тов либо снижается (менее 4,0 . 109), либо
повышается (более 8,0 . 109), то это признаки неполноценности,
напряженности реакции. Увеличение числа эозинофилов говорит о
глюкокортикоидной недостаточности: при развитии реакций активации и
тренировки — об относительной недостаточности, при развитии
стресса — абсолютной. Необходимо от-метить, что сочетание
эозинофилии с лимфопенией — неблагоприятный признак,
свиде-тельствующий о наступающем истощении глюкокортикоидной функции
коры надпочеч-ников. Увеличение числа лимфоцитов (более 40—45%
— в зависимости от индивидуаль-ных особенностей),
сопровождающееся нарушением хорошего самочувствия (появление каких-либо
жалоб или заболевания), свидетельствует о развитии переактивации.
Все эти отклонения являются сигналом для небольшого уменьшения дозы с
целью перевода данной реакции в рамки ее нормальных параметров.
При наличии признаков напряженности число сегментоядерных нейтрофилов
мо-жет не соответствовать величине, указанной для каждой реакции.
О методике взятия крови и подсчета формулы белой крови для определения
характера реакции. Предлагаемые сравнительно простые критерии оценки
общих адап-тационных реакций, развивающихся в организме при действии
неспецифических раздра-жителей, могут быть использованы для оценки
характера неспецифического действия раз-нообразных агентов и для
целенаправленной активационной терапии. Однако эти крите-рии позволяют
контролировать развитие адаптационной реакции лишь в случае соблюде-ния
следующих дополнительных условий:
1. Подсчет лейкоцитарной формулы должен проводиться не менее чем на 200 кле-ток, лучше на 300 клеток
137
(особенно при пограничных — между реакциями — значениях лимфоцитов).
2. Подсчет лейкоцитарной формулы должен проводиться по известному
способу: в 3—4 полях зрения (п. з.), по краю мазка, 3—4 п.
з. под прямым углом, 3—4 п. з. парал-лельно краю мазка, 3—4
п. з. под прямым углом по направлению к краю мазка и т. д. до набора
половины числа подсчитываемых клеток. Вторая половина подсчитывается
так же по другому краю мазка (Справочник по клиническим и лабораторным
методам исследова-ний. М., 1975. С. 46).
3. Подсчет в динамике у одного и того же больного должен осуществляться одним и тем же врачом-лаборантом.
4. Взятие крови должно производиться в одно и то же время суток,
желательно — утром и натощак, особенно при необходимости точного
учета числа лейкоцитов (лейко-пения, лейкоцитоз).
5. Взятие крови должно проводиться до лечебной процедуры (приема лекарств).
6. Взятие крови проводится: в день начала лечения (или накануне)
— исходный фон, через день после начала лечения и затем два раза
в неделю. При длительном лечении и хорошем состоянии больного анализы
можно проводить один раз в неделю.
Показания к использованию реакций тренировки и активации. Реакция
трени-ровки (в ее начальной стадии) используется для достижения
противовоспалительного эф-фекта (острые воспалительные процессы:
гастрит, панкреатит, колит, обострение язвен-ной болезни, аднексит,
бронхит, острый тромбофлебит, инфекционный неспецифический полиартрит),
для снижения свертывания крови, а также для защиты при повреждениях
ле-чебными воздействиями (лучевое лечение, химиотерапия), особенно
сопровождающимися выраженной лейкопенией.
Реакция активации применяется, напротив, в случаях, где воспалительный
потен-циал снижен, с целью его повышения, например, при хронических
вяло текущих воспали-тельных процессах (хронический гастрит,
панкреатит, колит, бронхит, неспецифическая пневмония, в том числе у
детей, язвенная болезнь желудка и 12-перст-ной кишки, аднек-сит и др.);
при гипертонической болезни и ишемической болезни сердца; при
гипотонии; при опухолевой болезни; при повреждениях лечебными
воздействиями (массивная проти-вотуберкулезная терапия, применение
больших доз антибиотиков, не сопровождающихся выраженной лейкопенией);
при психических заболеваниях (шизофрения, маниакально-депрессивный
психоз, реактивный психоз). Реакция повышенной активации (РПА) более
эффективна, но ее трудно поддерживать у больных. РПА также эффективна
для снижения коагуляционных свойств крови.
При активационной терапии воспалительных процессов реакции тренировки и
ак-тивации должны вызываться на сравнительно низких уровнях
реактивности: дозы лишь в 5—50 раз меньше терапевтических.
При использовании же реакции тренировки в каче-
138
стве защитного средства при повреждениях лечебными воздействиями,
особенно сопро-вождающимися выраженной лейкопенией, должны применяться
воздействия в дозах в 50 — 10000 раз меньших, чем
терапевтические, т. е. использоваться высокие уровни реак-тивности. То
же самое относится к реакции активации при лечении сердечнососудистых
заболеваний и опухолевой болезни (в большинстве случаев).
Прием биостимуляторов или введение нейротропных средств без
сознательного, целеустремленного вызова нужной для данного случая
адаптационной реакции на опреде-ленном уровне реактивности не является
эффективным. Для достижения лечебного эф-фекта необходимо получение и
стойкое поддержание либо реакции тренировки, либо ре-акции активации.
Исследования показали, что наиболее стойкой реакцией здоровья явля-ется
реакция повышенной активации, обусловливающая высокую неспецифическую
рези-стентность организма..
В прогностическом отношении наименее благоприятна стойкая стадия
истощения стресса. Развитие реакции тренировки благоприятно при наличии
показаний. Наиболее благоприятно в прогностическом отношении развитие и
длительное поддержание реакции активации, чаще всего активации на
высоких уровнях реактивности. Развитие реакции ак-тивации, особенно
РПА, обычно предшествует выздоровлению.
С характером развивающейся адаптационной реакции связаны течение и
прогноз патологического процесса. Целенаправленное использование
реакций тренировки и осо-бенно активации является основой активационной
неспецифической терапии. Одновре-менно с активационным неспецифическим
лечением может проводиться специальное ле-чение данного заболевания.
Противопоказаний к проведению активационной неспецифи-ческой терапии
пока не обнаружено.
Повторяемость адаптационных реакций. Понятие о коэффициенте реакции.
Необходимо учитывать, что четыре названные адаптационные реакции
повторяются на различных уровнях реактивности организма
(«этажах»).
Коэффициент реакции (К) — величина, на которую нужно умножить
дозу, чтобы перейти в следующую реакцию,— характерен для каждого
человека. Для практически здоровых К колеблется от 1,2 до 1,3, чаще
1,2; у больных ослабленных, пожилых К бывает и меньше: 1,15, 1,1.
Обычно, чем старше человек, или чем тяжелее его состояние, если это
больной, тем ниже К.
Таким образом, если методом проб (см. «Как подбирать дозу»)
выяснено, что ко-эффициент равен, например, 1,2, для перехода из
реакции тренировки в реакцию актива-ции надо увеличить дозу в 1,2
раза (т. е. на 20%), в реакцию стресс —
в 1,2X1,2 (т. е. на
139
44%). Для перехода в такую же реакцию следующего диапазона надо
увеличить дозу в 1,2X1,2X1,2X1,2, т. е. умножить на коэффициент в
четвертой степени (в два раза). Так же можно переходить на любой
уровень реактивности.
Таким образом, подбор доз сводится к нахождению коэффициента реакции в каж-дом отдельном случае.
Чем и как вызывать описанные реакции. Могут быть использованы различные
средства, способные вызывать адаптационные реакции путем воздействия на
организм через гипоталамо-гипофизарную систему (как непосредственно,
так и рефлекторно).
Установлена эффективность в этом отношении относительно нового
физиотерапев-тического средства — магнитного поля (МП). МП
действует дистантно, мягко, без повре-ждения. Постоянное МП вызывает
преимущественно реакцию тренировки на разных уровнях реактивности даже
без специального подбора «дозы». Переменное МП при под-боре
«дозы» (индукции, экспозиции, частоты или формы импульса)
может вызывать раз-личные адаптационные реакции. Учитывая
противовоспалительное действие реакции тре-нировки и максимальное
повышение резистентности при реакции активации, часто при-ходится
сочетать применение постоянного и переменного МП, особенно при
необходимо-сти сочетания общего и местного действия.
Используется ряд лекарственных нейротропных средств, особенно тех,
которые образуются в организме (адреналин, норадреналин) в
физиологических (а не в фармаколо-гических) дозах, а также
биостимуляторов растительного и животного происхождения.
Главным условием для достижения определенного эффекта, т. е. вызова
нужной адапта-ционной реакции любым из применяемых средств, является
подбор дозы. Учитывая осо-бую эффективность реакций тренировки и
активации высоких уровней реактивности, можно применять дозы,
составляющие 1/10, 1/20, 1/50, 1/100, 1/1000 и даже 1/10000
тера-певтической. Например: 1. Подкожные инъекции разведенных растворов
адреналина (1 мл раствора адреналина 1:1000 из ампулы разводится 9 мл
стерильного физиологического раствора, т. е. в 10 раз) и затем подбор
дозы (по анализу крови), начиная с инъекции 0,3—0,5 мл
разведенного раствора адреналина, один раз в первой половине дня. При
необхо-димости увеличения дозы можно 1 ампулу адреналина разводить 4 мл
физ. раствора, т. е. в 5 раз, а при необходимости уменьшения дозы 1
ампулу адреналина можно разводить (последовательно) в 100, 1000 и даже
10000 раз. Активационные дозы адреналина часто вызывают кратковременную
гипогликемию. При наличии признаков гипогликемии необ-ходимо выпить
стакан сладкого чая.
2. Прием внутрь теофедрина или дибазола (индивидуально подбор дозы, по
анализу кро-ви, начиная, с 1/4-1/3 таблетки 1 раз в день).
140
3. Прием внутрь экстракта элеутерококка, золотого корня, левзеи,
пантокрина (для нача-ла 15—20 капель
1 раз в день утром, затем подбор дозы: увеличивать до
терапевтиче-ской дозы, уменьшать до одной капли экстракта,
разведенного водой в 10 раз).
4. Магнитное поле, переменное, частотой
0,1—50 Гц, экспозицией 1 — 10 мин при
воз-действии на голову (в области затылка на уровне
прямой, проходящей через
ушные отверстия) индукцией 0,1 —10 мТ и
на патологический очаг — переменное или постоян-ное
магнитное поле — экспозицией
5—20 мин, индукцией
от 5,0 до 100 мТ.
Как подбирать дозу (по анализам крови).
1. Если после первого воздействия кровь становится характерной для
реакции тре-нировки то в случае, когда нужно поддерживать реакцию
тренировки, дозу оставить прежней (до следующего анализа). Если же
нужно вызвать реакцию активации, дозу уве-личить, умножив на
предполагаемый коэффициент реакции (К). Если после этого разо-вьется
реакция активации, значит К подобран правильно — дозу оставить
такой же (до следующего анализа). Если окажется, что К не соответствует
действительному, возможны два варианта: а) остается реакция тренировки
— тогда надо повторно увеличить первона-чальную дозу, взяв
большой коэффициент; б) вместо реакции активации разовьется реак-ция
стресс — коэффициент надо уменьшить.
2. Если после первого воздействия кровь становится
характерной для реакции ак-тивации, то в случаях, когда нужно
поддерживать реакцию активации, дозу оставить прежней. Если же
нужно вызвать реакцию тренировки, необходимо уменьшить дозу, раз-делив
на предполагаемый коэффициент реакции.
3. Если после первого воздействия кровь становится
характерной для стресса, то дозу нужно уменьшить до следующего анализа,
разделив на величину предполагаемого коэффициента. Если после
этого остается стресс, то выбранный
коэффициент слишком мал, если реакция тренировки —
слишком велик, если реакция активации — величина ко-эффициента
подобрана правильно.
4. При расчете коэффициента (К)
необходимо учитывать, что самое главное — не
попасть в стресс. Например:
а) если нужно перевести из реакции тренировки в
реакцию активации, то из двух предполагаемых коэффициентов
(если К данного челове-ка еще не определен), нужно выбрать меньший, так
как лучше остаться в реакции трени-ровки, чем перейти в стресс;
б) если нужно вывести из стресса в
реакцию активации, то из двух
предполагаемых К нужно брать наибольший, так как лучше
перейти в реак-цию тренировки, чем остаться в реакции стресс.
5. Необходимо помнить, что анализ может не измениться и
ввиду стойкости реак-ции, в которой до воздействия длительное время
находится организм, и
развития со-стояния ареактивности.
Кроме
141
стойкого состояния здоровья (РПА) имеется два стойких патологических
состояния: хро-нический стресс и переактивация. Поэтому в случае, когда
организм не удается вывести из стойкой реакции путем изменения дозы в
пределах одного уровня реактивности, целе-сообразно изменить ее так,
чтобы перейти на другой, вначале уменьшив для перевода на 1-2 и даже 3
уровня ниже, а затем (если и это окажется неэффективным) —
увеличить до-зу для перевода на 1 уровень выше первоначального.
6. Если вызванная нужная реакция на данном уровне
реактивности не оказывает
существенного лечебного эффекта, то
также необходимо перевести организм на
другой уровень реактивности (см. п. 5).
7. При наличии признаков напряженности для сохранения данной
реакции дозу надо немного уменьшить, разделив на vК, т. е.
чаще всего уменьшить на
10%. Для перевода в
вышележащую реакцию (например, из
напряженной тренировки в
ак-тивацию) дозу надо немного увеличить, умножив на - vК,
т. е. также чаще всего увели-чить на 10%. Для перевода в
нижележащую реакцию (например, из напряженной актива-ции — в
тренировку) уменьшить дозу несколько более обычного, разделив на К vК,
т. е. чаще всего уменьшить на 30%.
При повторных воздействиях принцип подбора дозы остается таким же. При
угрозе лейкопении дозу всегда необходимо уменьшить до получения реакции
тренировки (этого или предыдущего уровня).
В моменты подъема числа лейкоцитов свыше 5,0 . 109 дозу можно увеличить для периодического вызова реакции активации.
8. В промежутках между анализами дозу (силу) действующего
фактора лучше не-много (на 5—10%) менять или в виде
«зубчика» или небольшой волны.
Кроме соотношения форменных элементов белой крови помочь в подборе дозы может субъективное состояние больных.
Для реакции тренировки характерно спокойствие, небольшая сонливость,
иногда легкое кратковременное головокружение (пошатывание), хорошее
самочувствие, сон, ап-петит.
Для реакции спокойной активации характерно спокойное, бодрое состояние,
хоро-ший сон, аппетит. Для реакции повышенной активации характерно
приподнятое настрое-ние, даже иногда с оттенком эйфории, жажда
деятельности, хороший сон, отличный аппе-тит.
При наличии выраженных элементов напряженности (развитие реакций на
низких уровнях реактивности) появляются признаки нарушения самочувствия
(раздражитель-ность, апатия, нарушения сна, плохое настроение) даже при
реакциях тренировки, РСА и РПА.
Примечания.
1. Если исходный анализ характерен для активации, это не является
противопока-занием для специального поддержания
длительной активации. Периодическая
акти-вация может
142
возникнуть даже среди болезни, но она не бывает стойкой и большей
частью бывает не-полноценной или развивается на одном из высоких
«этажей».
2. Если отмечается переактивация (число
лимфоцитов больше 45%), это плохой признак, особенно
в сочетании с лейкопенией. Такое состояние нужно
изменять, пере-водя в рамки реакции активации или даже тренировки.
3. При хронической пневмонии у детей соотношение форменных
элементов в лей-коцитарной формуле характерно для стресса взрослых. При
активационной терапии жела-тельно получить РПА (также по параметрам,
описанным для взрослых).
4. Необходимо учитывать индивидуальные особенности каждого
человека, так как для разных людей оптимален свой уровень
активации (уровень числа лимфоцитов, при котором
отмечается хорошее самочувствие). Поэтому верхней границей РПА по числу
лимфоцитов считаем 40—45%.
Как видно из методических рекомендаций, подбор дозы по лейкоцитарной формуле применяется при различных заболеваниях.
Для автоматического расчета доз по лейкограмме нами предложено
специальное устройство (А. с. № 754726 на имя И. Г. Коп-па, Н. И.
Пинчука, Л. X. Гаркави и Е. Б. Ква-киной).
С профилактической (а также лечебной) целью в «третьем
состоянии», состоянии предболезни и болезни (если нельзя часто
брать кровь на анализ) можно применять про-граммированные режимы вызова
и поддерживания реакции активации высоких уровней реактивности, не
требующие частого исследования крови.
О программированных режимах и их месте
в активационных оздоровлении, профилактике и лечении
На основании многолетнего исследования закономерностей развития
адаптацион-ных реакций на разных уровнях реактивности оказалось
возможным разработать про-граммированные режимы дозировки
неспецифических средств и воздействий для вызова и длительного
поддержания в организме оптимального состояния здоровья — реакции
ак-тивации (в первую очередь, повышенной) без постоянного использования
обратной связи с организмом по лейкограмме, как это делалось ранее.
Каждый режим имеет свои особенности — наибольшую эффективность по
отношению к определенной категории людей, оптимальные сроки применения,
границы дозовых и силовых характеристик и т. п., но все режимы
отличаются простотой в исполнении, доступностью для каждого,
надежностью и высокой эффективностью. Можно сказать, что за исключением
самых тяжелых заболева-ний (например, поздних стадий опухолевых
болезней) эти режимы,
если их исполь-зовать
правильно, работают
143
не хуже, чем подбор дозы но обратной связи. Более того, когда в тяжелых
случаях при стойком высокоэтажном стрессе или переактивации не удается
перевести организм в фи-зиологическую реакцию подбором дозы по обратной
связи, мы часто рекомендуем на ко-роткое время определенный
программированный режим, который нацелен именно на вы-ведение из
стойкого состояния (напри мер, стрессорной ареактивности), а затем
снова пе-реходим на подбор дозы по обратной связи. Между курсами
специального лечения, когда анализы крови брать затруднительно, также
целесообразно пользоваться программиро-ванными режимами. Но самая
большая область применения программированных режимов — это
оздоровление и профилактика по отношению ко всем возрастным категориям
лю-дей, т. е. фактически ко всем, кто еще не болен, так как вряд ли
целесообразно рекомендо-вать практически здоровым людям систематический
контроль за характером адаптацион-ной реакции по анализу крови (да это
и нереально).
Вторая — также очень большая область применения этих режимов
— это лечение начальных стадий самых различных заболеваний, когда
еще нет грубых структурных на-рушений.
Третья область — это лечение старения, поскольку и у этой
категории людей часто брать анализы сложно, а эффективность режимов при
этом не ниже.
И, наконец, четвертая область — это применение программированных
режимов для улучшения состояния людей с постоянными большими
психоэмоциональными или физи-ческими нагрузками и людей, постоянно
подвергавшихся действию повреждающих факторов.
Программированный режим № 1 основан на использовании принципов новизны
и минимизации воздействий (разработан совместно с Л. Н. Лупичевым и Е.
А. Файдышем (Институт физико-технических проблем, г. Москва)).
Известно, что при развитии реакции активации, наиболее повышающей
сопротивляемость организма, в мозгу преобладает умеренное возбуждение.
Преобладание возбуждения развивается в мозгу в ответ на но-визну
действующего фактора. Новизна может быть связана с качеством
раздражителя (спецификой) и с количеством. Для того, чтобы
«фактор новизны» приводил к преобладанию не чрезмерного, а
физиологического возбуждения, новизна также должна быть не чрезмерной,
т. е. принцип новизны должен сочетаться с принципом минимизации
воз-действия.
Для поддержания хорошего уровня здоровья программированный режим № 1 мож-но использовать долго, месяц и более.
Применение режима № 1 разработано с учетом пола, возраста, исходного состоя-ния и характера имеющихся заболеваний.
Режим № 1 может быть применен:
1) у здоровых, для поддержания уровня здоровья;
144
2) в состоянии предболезни
(донозологическое состояние) без четко выражен-ных местных
проявлений — профилактическое лечение. Такое лечение особенно
показано весной и осенью, когда частота заболеваний увеличена;
3) параллельно с проведением специального
лечения (в поликлинике и при не очень тяжелых
заболеваниях в стационаре) для создания хорошего неспецифического
фона, что повышает эффективность специфического лечения;
4) в промежутках между проведением
курсов специфического лечения для пре-дупреждения рецидивов
хронических заболеваний и поддержания здоровья.
Результаты применения режима № 1 показали его высокую эффективность для
оз-доровления, профилактики и повышения эффективности специального
лечения. Так, за-болеваемость у практически здоровых людей (более 500
человек) после двукратного, в течение 1 — 1,5 месяцев применения
режима № 1 снизилась на 30—50%. Заболевания, если они возникали,
протекали легче и выздоровление наступало быстрее примерно на 1/3 срока
заболеваний в контрольной группе. Исследование лейкограммы, как
простого пока-зателя реакции, показало, что исходные анализы
свидетельствовали о наличии, в основ-ном, реакций активации и
тренировки с небольшими элементами напряженности и иногда —
мягкого стресса. После применения режима стресс почти не встречался и
элементы на-пряженности реакций отмечались крайне редко.
Применение режима № 1 у людей с донозологическими состояниями (более
300 человек) — с жалобами на общую слабость, быструю
утомляемость, частые головные бо-ли, боли в области сердца, плохой сон
и т. д. и т. п.— показало, что после применения ре-жима № 1 в
течение 1-2-х месяцев жалобы исчезали в 80% случаев, а в 15% —
станови-лись менее выраженными. В тех случаях, когда состояние
существенно не изменялось, обычно обнаруживалось наличие какого-либо
заболевания.
Таким образом, режим оказался высоко эффективным для лечения
донозологиче-ских состояний, что, учитывая доступность его для каждого
и дешевизну, открывает пер-спективы активной профилактики широких масс
людей.
Анализы крови до начала применения режима № 1 свидетельствовали о
преоблада-нии неглубокого стресса и реакций активации и тренировки с
элементами напряженности, более выраженными, чем в группе практически
здоровых людей. После применения режима лейкограммы в большинстве
случаев нормализовались: преобладали реакции акти-вации и тренировки
без выраженных элементов напряженности. При использовании ре-жима № 1
параллельно с проведением специального лечения (в поликлинике и в
стационаре) у более 250 человек отмечалось повышение лечебного эффекта,
145
более быстрое выздоровление (также примерно на 1/3 срока по сравнению с
контрольной группой), более быстрое исчезновение симптомов болезни при
хорошем общем состоя-нии.
Если режим № 1 принимался и между курсами специального лечения, то
рецидивы хронических заболеваний возникали реже и протекали легче. При
этом общее состояние организма нарушалось существенно меньше.
Программированный режим № 2 активационной терапии основан на
использова-нии биоритмов реакций и разной чувствительности на разных
иерархических уровнях ор-ганизма. Применяется при наличии исходного
стойкого состояния предболезни и болезни, для восстановления физической
и умственной работоспособности, особенно при наличии выраженного
местного патологического процесса, для выведения из патологического
со-стояния, в том числе стойкого, при необходимости сочетания общего и
местного дейст-вия. Режим может быть использован в физиотерапии и
рефлексотерапии. Применение ре-жима № 2 более чем у 200 человек с
различными хроническими заболеваниями, в том числе тяжелыми (вплоть до
онкологических), показало его высокую эффективность (выше 80%).
Сочетание при этом общего и местного действия приводило к быстрому
улучшению местного патологического процесса с исчезновением или
существенным уменьшением жалоб. Полное выздоровление наступало при
излечимых в настоящее время заболеваниях. В других случаях отмечалось
существенное улучшение и повышение эффективности спе-циальных методов
лечения. Высокая эффективность режима № 2 открывает перспективу его
широкого использования для повышения эффективности лечения простым и
экономичным путем.
Исследование лейкограммы в исходном состоянии выявляло наличие либо
глубо-кого стресса, либо реакций тренировки и активации с несколькими и
резко выраженными признаками напряженности. При выздоровлении
развивалась, как правило, реакция активации с незначительно выраженными
признаками напряженности. При улучшении со-стояния (разной степени)
отмечались либо стресс, значительно более мягкий, чем исход-ный, либо
реакции тренировки и активации с меньшим числом и выраженностью
призна-ков напряженности.
На программированный режим № 2 получено авторское свидетельство №
1474925 (на имя Л. X. Гаркави, Е. Б. Квакиной, Л. Н. Лупичева, Е. А.
Файдыша, Т. С. Кузьменко).
Программированный режим № 3 основан на использовании экспоненциальной
зависимо-сти между дозой (силой) действующего фактора, а также принципа
минимизации. Уже давно было замечено наличие экспоненциальной
зависимости в динамике адаптационного процесса
(Adolph E. F., 1956).
Многолетние исследования, как
146
наши, так и Ю. В. Серегина (1986), показали в определенных случаях
наличие экспонен-циальной зависимости между величиной действующего
фактора и характером ответной адаптационной реакции. Эта зависимость
наиболее часто используется нами при наличии стойких патологических
состояний. К ним мы относим стресс низких уровней реактивно-сти
(развивающийся в ответ на длительное воздействие большого по абсолютной
величи-не действующего фактора, большей частью — патологического
процесса) и переактива-цию — напряженную по лимфоцитам
(лимфоцитоз) повышенную активацию низких уровней реактивности
(развивающуюся в ответ на длительное действие больших нагрузок, чаще у
молодых, сохраняющих длительно видимость здоровья). Кроме того, иногда
таки-ми состояниями являются реакции тренировки и активации низких
уровней реактивности с резко выраженными и стойкими элементами
напряженности.
Для всех этих состояний типично отсутствие изменений характера
адаптационной реакции на применение малых и даже не очень малых
воздействий как по обратной связи с организмом (по лейкоцитарной
формуле), так и по программированным режимам № 1 и № 2. Применение же
режима № 3 оказалось высоко эффективным для выведения орга-низма из
таких состояний и возвращения к норме реакции. Наиболее эффективен
экспо-ненциальный режим и для преодоления стойкой лейкопении.
Использование режима № 3 при различных заболеваниях и побочном влиянии
ле-чения при действии токсических химиопрепаратов и облучения показало
его высокую эффективность. С помощью этого режима удалось вывести из
стойких патологических состояний (как в результате болезни, так и
действия длительных перегрузок и повреж-дающих факторов) более 300
человек.
При этом отмечалось улучшение общего самочувствия, существенное
уменьшение жалоб, исчезновение или существенное уменьшение лейкопении,
увеличивалась эффек-тивность специального лечения. Если до начала
применения режима № 3 лейкограмма свидетельствовала о преобладании
тяжелого стресса, переактивации или реакции трени-ровки и активации с
резко выраженными признаками напряженности, то после его прове-дения
отмечались нормальные реакции тренировки и активации или эти же
реакции, но с небольшими признаками напряжения и неглубокий стресс.
Показания к применению программированных режимов активационной
про-филактики и терапии. Применение того или иного программированного
режима зависит от характера исходного состояния и, прежде всего, от
того, насколько оно стойкое.
Если функциональное состояние организма не отличается высокой
устойчивостью (кроме стойкого состояния здоровья), то целесообразно для
вызова и поддержания реакции акти-вации на физиологических
147
(высоких) уровнях реактивности применять программированный режим № 1.
Если имеется стойкое патологическое состояние, например, хронический
стресс, переактивация или реакции тренировки и активации с резко
выраженными и стойкими элементами напряженности, то в случаях, когда
оно сочетается с выраженным местным патологическим процессом, лучше
применять сначала программированный режим № 2. Если же при стойком
состоянии местный очаг не выявлен или режим № 2 оказался
мало-эффективным, можно начинать с другого программированного режима
— № 3. Как в слу-чае применения режима № 2, так и режима № 3,
через 2—4 недели можно переходить на программированный режим № 1.
Как определить, является ли состояние стойким? Для этого необходимо
исследо-вать лейкоцитарную формулу и общее число лейкоцитов 2 — 3
раза с интервалом 2—3 — 5 дней. Если лейкограмма от анализа
к анализу существенно (более, чем ошибка счета) меняется (даже в
пределах одной реакции), а не остается одинаковой, то состояние
не-стойкое, и оно подлежит лечению по программированному режиму № 1.
Если речь идет о курортном лечении, то для суждения о том, стойкое
состояние или нет, достаточно двух анализов: сразу же после приезда и
через 3—4 дня, так как сам переезд является своеоб-разной
нагрузочной функциональной пробой. Если лейкограмма не меняется,
состояние стойкое. Стойкие состояния отмечаются при переактивации
(процентное содержание лимфоцитов более 40—45%), с нарушениями
самочувствия, в первую очередь — сна; хроническом стрессе
(процентное содержание лимфоцитов менее 20%) и иногда — при
реакциях тренировки и активации высоких этажей, когда имеются резко
выраженные и также стойкие признаки напряженности реакций: моноцитоз
или монопения, эозинофилия или анэозинофилия, лейкопения или
лейкоцитоз, базофилия.
Если есть возможность, через 3—4 недели проведения режима
желательно вновь исследовать лейкоцитарную формулу, чтобы определить,
повторить ли режим № 3 или переходить на режим № 1. Если исходный
анализ существенно не изменился — организм не вышел в норму
реакции, режим № 3 можно повторить и лишь после этого переходить на
режим подбора дозы по режиму № 1.
Для профилактики и лечения хронической лучевой болезни и поздних
лучевых по-вреждений (например, при лучевом лечении онкологических
больных), а также для по-вышения неспецифической резистентности,
оздоровления организма у населения в регионах, подвергающихся
(подвергавшихся) действию радиации, могут быть использованы режимы: № 3
— при наличии лейкопении, а когда число лейкоцитов нормально
— режим № 1.
Даже в случаях, если действие радиации не было точно установлено,
148
предлагаемые нами методы повышения неспецифической резистентности будут
способст-вовать повышению уровня здоровья. Необходимо подчеркнуть, что
предлагаемый способ не испытывался по отношению к массивному
(однократному) действию ионизирующей радиации. Что же касается его
применения при длительном (повторном) действии радиа-ции в сравнительно
небольших дозах, но превышающих допустимые, оно может осущест-вляться
как самостоятельно, так и совместно со специальными средствами
биологической защиты (радиопротекторами), что повышает эффективность
последних.
В разработанных программированных режимах предлагается использовать для
вы-зова и поддержания реакций тренировки и активации самые различные
биостимуляторы растительного и животного происхождения, в том числе
широко известный препарат природного (растительного) происхождения
— элеутерококк, который в терапевтических до-зах применяется для
повышения сопротивляемости при различных заболеваниях и, в том числе, в
комплексной терапии лучевой болезни (Брехман И. И., 1982). Однако при
традиционном применении не учитывается влияние на характер общей
адаптационной реакции, а именно это влияние, как уже говорилось,
обусловливает уровень резистентности орга-низма, т. е. определяет во
многом эффективность действия препарата.
В предлагаемых режимах с помощью адаптогена (элеутерококка) в организме
це-ленаправленно вызывается нужная адаптационная реакция, что не только
значительно по-вышает его эффективность, но и позволяет использовать
лекарственный препарат в значи-тельно меньших дозировках, что полезно
для ослабленного повреждающим фактором ор-ганизма и, кроме того, в
условиях массового употребления высоко экономично.
Хочется подчеркнуть, что программированные режимы можно применять во
многих ва-риантах и сочетать, например, вначале режим № 1 с режимом № 2
для вывода из неблаго-приятного состояния, а затем перейти к режиму № 1
для поддержания хорошего состоя-ния. Таким образом, применение
программированных режимов при учете пола, возраста, индивидуальных
особенностей, характера и тяжести нагрузок или заболеваний открывает
перспективы дальнейших исследований с целью совершенствования подходов
к управле-нию резистентностью организма.
Отсутствие какого-либо побочного повреждающего действия и наличие той
или иной меры повышения степени здоровья позволили О. П. Щепину, Г. И.
Царегородцеву и В. Г. Ерохину рекомендовать наш подход для широкого
использования в практике дис-пансеризации населения (Медицина и
общество. М., 1983. С. 151).
149
Применение активационной терапии для
профилактики и оздоровления
Еще Гален подразделял состояние человеческого тела на три категории:
здоровье, болезнь и некоторое третье, промежуточное состояние —
ни здоровье, ни болезнь. Ави-ценна различал шесть степеней здоровья и
болезней, из которых только две относятся к болезни: «Бывает тело
здоровое до предела, тело здоровое, но не до предела, тело не
здо-ровое, но не больное... затем тело в хорошем состоянии, быстро
воспринимающее здоро-вье, затем тело, больное легким недугом, затем
тело, больное до предела». Не останавливаясь далее на вопросах
классификации, мы хотим лишь обратить внимание на тот факт, что
«третье состояние» — состояние предболезни,
промежуточное между здоровьем и бо-лезнью, встречается очень часто
(Баевский Р. М., Казначеев В. П., 1974; Брехман И. И , 1976).
В настоящее время в медицине большое значение придается
«активно-конструктивной» (наступательной) профилактике, в
которой особо выделяется «формиро-вание или усиление уже
имеющихся оздоравливающих механизмов жизнедеятельности организма»
(Щепин О. П., Царегородцев Г. И., Ерохин В. Г., 1982). Для
осуществления такой активной профилактики нужно уметь выявлять не
тяжело больных, а людей с не-большими отклонениями от состояния
здоровья — в донозологических состояниях или в начальных стадиях
заболевания, когда имеются еще лишь функциональные нарушения. После
выявления их нужно лечить, возвращая к состоянию здоровья, причем по
возмож-ности без применения сильнодействующих лекарств (Ромашов Ф. И.,
Фролов В. А., 1983; Кацман М. В., Сидоренко Ю. С, Айзенштарк Э. А.,
1986).
Крайне важный принцип активной профилактики и лечения — принцип
этапности. Первый этап — повышение уровня здоровья, который, по
нашему мнению, основанному на многолетних исследованиях, нужно
проводить с юности. В. М. Дильман (1987) считает, что активную
профилактику — профилактическое лечение — нужно начинать с
20—25 лет, когда заканчивается рост и развитие организма,
необходимые для воспроизведения потомства, и «закон отклонения
гомеостаза» начинает работать на формирование главных
неинфекционных болезней и старения. Мы думаем, что иногда, например,
если ребенок часто болеет, курсы «профилактического
лечения» — неспецифического, повышающего резистентность
организма, с помощью использования очень малых доз биостимуляторов
можно (и нужно) проводить с детства.
Какое значение активационная терапия и профилактика могут иметь для
самых здоровых людей, которые, согласно Авиценне, должны достигать
предела здоровья, на-пример, для спортсменов?
150
Спортивные тренировки нацелены на то, чтобы были получены наилучшие
результаты в своем виде спорта. Исходя из наших представлений, хорошей
спортивной форме спорт-сменов должна соответствовать реакция активации,
а в период соревнований должна быть преобладающей реакция повышенной
активации, при которой человек ощущает эмоцио-нальный подъем, большой
прилив энергии: он как бы способен сделать больше того, что может
обычно. Особенно важно то, что это дается не ценой разрушения организма
(как при стрессе или при приеме допингов), а физиологическим путем как
по характеру изме-нений в регуляторных и защитных системах организма,
так и по энергетике. Поэтому для режима спортивных тренировок
необходимо учитывать не только характер развивающейся адаптационной
реакции, но и уровень реактивности. Перетренировки сопровождаются часто
развитием стресса или активацией на чрезмерно высоких этажах —
переактивацией, что может привести к срыву.
По нашему мнению, для предотвращения срыва или для выхода из него
необходи-мо поддерживать в организме реакцию активации на низких
этажах, например, с помо-щью малых доз какого-либо биостимулятора, что
минимизирует энергозатраты организма. Если спортсмен из
«состояния здоровья» переходит в «третье
состояние», то он должен подлежать неспецифическому лечению
согласно описанным выше правилам. Если спорт-смен переходит в категорию
«вообще больных», то целенаправленное развитие показан-ной
в каждом случае определенной адаптационной реакции могло бы оказать
большую помощь специализированному лечению, будь то оперативное
вмешательство или консер-вативные лечебные мероприятия. Известно, что
наши чемпионы получают высококвали-фицированное специальное лечение
(Миронова 3. С., 1976). Однако зачастую трудность лечения, длительность
заболевания и неполное восстановление функций связаны с тем, что общий
неспецифический фон не препятствует, а способствует развитию
заболевания. В этих случаях особенно необходимо целенаправленное
развитие определенной адаптационной реакции.
В отношении еще одной категории «здоровых до предела» людей
— космонавтов — тоже хочется отметить целесообразность
учета состояния организма не только с помо-щью высокоспециализированных
методов анализа работы его различных систем, но и с помощью
установления характера общей адаптационной реакции. В. В. Парин и Р. М.
Ба-евский (1970) указывали на возможность изучать «влияние
внешних условий на внутрен-нюю среду организма по автономным ответам
отдельных органов или изменениям от-дельных функций. В целостном
организме любая реакция осуществляется согласованным ансамблем
разнообразных систем». Мы думаем, что повышение неспецифической
рези-стентности с по-
151
мощью развития общих адаптационных неспецифических реакций целого
организма — реакций тренировки и особенно активации —
найдет свое место в системе тренировок космонавтов, тем более, что
исследованиями В. В. Парина, П. В. Васильева, В. Е. Белай (1965)
показано, что для повышения устойчивости к нагрузкам с помощью
фармакологи-ческих средств необходимо подбирать дозу не слишком
большую, но и не слишком ма-лую.
По мнению Парина и Баевского (1970), одной из важнейших задач
современной физиологии является разработка критериев для выявления
самых минимальных измене-ний в организме, возникающих при
субэкстремальных воздействиях. Эти критерии в дальнейшем могут стать
основой врачебного контроля и профилактики. Поэтому мы на-деемся, что
показатели реакций тренировки и активации, являющихся разными
варианта-ми нормы, войдут в комплекс критериев для оценки эффективности
медицинской подго-товки космонавтов к полету, тем более, что вопросы
адаптации и стресса стоят в центре внимания космической физиологии
(Газенко О. Г., Генин А. М., 1976).
Необходимо отметить особую роль движения, мышечной нагрузки для
успешного проведения активационной профилактики и терапии. О вредном
влиянии гипокинезии и гиподинамии и, напротив, о пользе двигательной
мышечной нагрузки говорят буквально все. Очень интересны работы И. А.
Аршавского (1968, 1979), который обнаружил «энер-гетическое
правило скелетных мышц». Смысл этого правила в том, что
двигательная ак-тивность является фактором функциональной индукции
анаболизма, цель которого не просто восстановление исходного состояния,
а избыточное восстановление. Избыточный же анаболизм — форма
негэнтропии, свойственной живому организму как открытой раз-вивающейся
системе. Мы пришли к такому же выводу на основании представлений о
ро-ли синхронизации для формирования адаптационных реакций, особенно
реакции актива-ции высоких уровней реактивности — неспецифической
основы здоровья. Движение, мышечную деятельность, особенно ритмическую,
мы рассматриваем как мощный и мяг-кий синхронизатор работы подсистем
организма. Поэтому при проведении активационной профилактики и терапии
на всех этапах необходимо сочетание действующих факторов с двигательной
активностью, особенно ритмического характера. Конечно, величина этой
активности на разных этапах должна быть различной и ни в коем случае не
чрезмерной, утомительной.
Необходимо подчеркнуть, что применительно к любым действующим факторам,
используемым для активационной профилактики и терапии, всегда
учитывается принцип минимизации — полезности постепенного
уменьшения их абсолютной величины для пе-рехода на более физиологичные
высокие уровни реактивности. Двигательная
152
активность является исключением. Ее необходимо постепенно, нелинейно
увеличивать до оптимального уровня — соблюдение принципа
оптимальной индивидуальной физической нагрузки (Меделяновский А. Н.,
1981; Меделяновский А. Н. и др., 1985). При таком по-вышении
двигательной активности до оптимальной не происходит нежелательного
пере-хода на более неблагоприятные, низкие уровни реактивности.
По-видимому, это связано с особой ролью двигательной активности в
метаболизме и синхронизационной деятельно-сти подсистем организма.
Очень важным для повышения уровня здоровья является второй этап
активной профилактики — лечение состояний, промежуточных между
здоровьем и болезнью. Мы сделали попытку рассмотреть эту категорию
людей с позиции теории адаптационных ре-акций.
Исследование людей, не относящихся к больным, но и не чувствующих себя
здоро-выми, показало, что большинство из них (примерно 80%) находится в
состоянии хрониче-ского стресса и значительно меньше (примерно 15%)
— в напряженных реакциях тренировки и активации. Так, например,
при числе лимфоцитов, характерном для реакции тре-нировки, может
наблюдаться эозинофилия, или реже — анэозинофилия, моноцитоз,
лей-копения или, напротив, лейкоцитоз. Те же самые нарушения могут быть
при числе лимфоцитов, характерном для реакции активации, с той
разницей, что здесь чаще наблюдается анэозинофилия. Кроме того,
отклонением от нормальной реакции активации является пе-реактивация,
когда число лимфоцитов выше 45%. Переактивация также часто
сопровождается анэозинофилией, лейкопенией или (редко) лейкоцитозом.
У совсем небольшой части обследуемых (около 5%) отмечались изменения,
укла-дывающиеся в картину обычной реакции тренировки или активации. Мы
могли лишь предположить, что в этих случаях адаптационные реакции
развивались на высоких эта-жах, когда они протекают с большими
энергетическими тратами и предъявляют к орга-низму большие требования.
Мы предприняли попытку лечения этих лиц. Применяли различные
биостимулято-ры, переменное магнитное поле (аппарат
«По-люс-1» и ГИМП), дозированную физиче-скую нагрузку и
водные процедуры. Кроме того, использовали соли янтарной кислоты
— сукцинат натрия и калия как биологически активные вещества,
играющие особую роль в энергетическом обмене (Кондрашова М. Н., 1972,
1974; Терапевтическое действие янтар-ной кислоты. Пущино, 1977). Мы
стремились вызывать и поддерживать реакцию активации на низких
«этажах» с помощью малой силы (дозы) воздействия, используя
мумие в дозе 5—50 мг, элеутерококк колючий — от 2 до 25
капель, сукцинат натрия или калия —0,25—0,5 г, переменное
магнитное поле с индукцией от 1 до 10 мТ в течение 2 — 5 мин. Все
эти воздействия (включая прием лекарств) применяли
153
ежедневно, один раз в день, в первую половину дня. Дозу воздействия для
получения ре-акции активации подбирали и корригировали либо по обратной
связи с организмом, либо по программированным режимам.
В тех случаях, когда до начала лечения у людей в «третьем
состоянии» отмечался хронический стресс, поддерживание у них не
только реакции активации, но даже и реак-ции тренировки приводило к
улучшению состояния, повышению работоспособности, ис-чезновению
неприятных ощущений, улучшению сна и аппетита, т. е. фактически
перево-дило их в состояние здоровья.
В случаях, когда до лечения были напряженные реакции тренировки или
актива-ции, нормализация реакций также переводила этих людей в
категорию здоровых. И, нако-нец, когда до лечения казалось бы были
нормальные реакции тренировки и активации, поддерживание этих реакций,
но заведомо на низких этажах (с помощью малых доз), име-ло такой же
лечебный эффект. Это говорит в пользу справедливости предположения о
том, что в такой группе плохое состояние было связано с развитием
реакции тренировки и активации на чрезмерно высоких этажах.
Преимущество низких этажей настолько суще-ственно, что применение малых
доз даже без контроля за развивающейся реакцией может дать неплохой
эффект при массовых профилактических мероприятиях.
Во время состояния «предповреждения»,
«предболезни» наблюдается нарушение гомеостаза (Чернух А.
М., 1976), поэтому целенаправленное развитие реакций, нормали-зующих
гомеостаз, особенно реакции повышенной и спокойной активации, должно
способствовать предотвращению развития, болезней. В. П. Казначеев
(1973) говорит о необ-ходимости специального изучения и лечения
предболезни, или «донозологического» со-стояния.
Если учесть, что в состоянии, промежуточном между здоровьем и болезнью,
нахо-дится большое число людей и что оно может длиться долго и чаще
всего переходит в бо-лезни, перевод этого состояния в состояние
здоровья есть одна из важнейших задач про-филактической медицины. Если
с помощью таких простых, доступных и щадящих орга-низм средств, вызывая
целенаправленно реакции тренировки и особенно повышенной и спокойной
активации, можно оздоравливать людей, находящихся в «третьем
состоянии», то, нам кажется, что этот метод целесообразно широко
внедрить в работу всевозможных профилакториев и различных учреждений
санаторно-курортного типа.
Хотя в санаториях и проводится специализированное лечение различных
заболева-ний, но каждому ясно, что специальное лечение не хуже, а может
быть, даже и лучше проводить в специализированных лечебных учреждениях.
Следовательно, основное, что выгодно отличает
санаторно-курортное лечение от
любого другого,— это
154
неспецифическое оздоравливающее действие различных курортных факторов:
климата, диеты, отдыха, развлечения, прогулок но терренкурам и т. д.
Однако использование этих оздоравливающих курортных факторов
нормируется, в лучшем случае, по их действию на отдельные системы
организма, без специального дозирования но общей ответной реакции.
Ознакомление с историями болезни в различных санаториях показало, что
лишь у не-большой части больных во время пребывания в санатории стойко
держится реакция акти-вации или тренировки. У остальных происходит
чередование различных реакций, в том числе часто встречается стресс. У
некоторых стресс является преобладающей реакцией. Таким образом, даже
при правильном проведении специального лечения основная задача
санаторно-курортного лечения - оздоровить человека, повысить его
устойчивость, сопро-тивляемость — выполняется далеко не всегда.
Действительно, если часть людей после са-наторно-курортного лечения
получает заряд на весь год, то у многих существенного, дли-тельного
улучшения не отмечается, а у некоторых даже наблюдается ухудшение. Мы
свя-зываем это именно с тем, что санаторно-курортное лечение не
использует целенаправлен-ного повышения резистентности.
Когда имеются жалобы общего характера, еще не указывающие на характер и
ло-кализацию развивающегося заболевания, требуется применение только
программирован-ных режимов. При лечении предболезни, донозологических
состояний, когда, кроме жа-лоб общего характера, появляются жалобы,
указывающие на конкретный характер и лока-лизацию развивающегося
заболевания, должно быть использовано и местное воздействие (например,
в режиме двойного воздействия), и, возможно, средства специального,
специ-фического лечения данного заболевания. И, наконец, третий этап
— лечение уже развив-шихся болезней. На него тратятся основные
силы и средства современной медицины. А между тем перенесение акцента
на категорию «здоровье», а не на категорию
«болезни», значительно снизило бы долю тяжелых заболеваний.
На этом этапе активационная тера-пия может применяться в комплексе со
специальной, специфической терапией, создавая в организме благоприятный
неспецифический фон, на котором специфическая терапия бу-дет более
эффективна.
При оперативном вмешательстве, когда даже несмотря на самую высокую
технику операции и хорошую психологическую подготовку пациента
развивается стресс, создание активационного фона до операции и
поддержание его в послеоперационный период зна-чительно ускоряет
выздоровление.
155
О принципе диагностического лечения
с целью выявления групп риска
и лечения заболевания на начальных стадиях
Очень сложным вопросом является выявление так называемых «групп
риска». Ис-пользуя активационную терапию, мы выдвинули принцип
диагностического лечения, со-единяющего в себе лечение начальных стадий
различных заболеваний и выделение групп риска.
Для своевременного обнаружения заболеваний в настоящее время
применяются различные способы диагностики, но во всех случаях
диагностика отделена от лечения. Однако «люди заинтересованы не в
том, чтобы их все более полно обследовали, а в том, чтобы их
своевременно и эффективно лечили и, главное, помогали бы им оставаться
здо-ровыми» (Царегородцев Г. И., Ерохин В. Г., 1986). Многие
способы выявления заболева-ний, особенно массовые профилактические
осмотры, анкеты, основываются на наличии определенных жалоб. Вместе с
тем общеизвестно, что у больных в начальных стадиях за-болевания, при
функциональных нарушениях субъективные неприятные ощущения выра-жены
даже ярче, чем при хронических заболеваниях. Дорогостоящие и
небезразличные для организма диагностические исследования на начальных
стадиях заболевания, при функциональных нарушениях мало информативны.
Многочисленные исследования показали, что активационная терапия, т. е.
исполь-зование специального вызова физиологических, неспецифических
адаптационных реак-ций организма — тренировки и особенно
активации — достигает эффекта наиболее быст-ро в донозологических
состояниях и на ранних этапах самых различных заболеваний. При тяжелых,
далеко зашедших заболеваниях с глубокими структурными нарушениями с
по-мощью активационной терапии эффект достигается медленно, и она
применяется, чаще всего, совместно с разработанной специфической
терапией данного заболевания, создавая для нее благоприятный
неспецифический фон. Это понятно, так как активационная тера-пия лишь
повышает неспецифическую резистентность, вследствие чего организм более
успешно борется со многими заболеваниями (Гаркави Л. X., Квакина Е. Б.,
1984; Гаркави Л. X., Квакина Е. Б., Уколова М. А., 1979). На начальных
стадиях многих заболеваний (га-стриты, холециститы, колиты, бронхиты и
т. д.), в донозологическом состоянии, предбо-лезни этого оказывается
достаточным для выздоровления.
Такая существенная разница в эффективности активационной терапии при
началь-ных и далеко зашедших стадиях различных заболеваний привела нас
к мысли об исполь-зовании ее в качестве диагностического лечения для
выявления групп риска и одновременного лечения донозологических
состояний, начальных стадий заболеваний и функ-циональных нарушений.
156
Для вызова и поддержания реакции активации без систематического
исследования лейкоцитарной формулы для коррекции дозы нами при
диагностическом лечении исполь-зовались программированные режимы.
Вначале удобно применять анкеты с перечнем основных жалоб, характерных
для определенных групп заболеваний, например, анкету, разработанную Э.
А. Айзенштарком для выявления заболеваний желудка и т. п. Затем в
течение месяца всем людям, предъяв-ляющим жалобы, проводится
активационная терапия и затем повторное анкетирование. Люди,
предъявляющие прежние жалобы, должны быть детально обследованы, они
со-ставляют группу риска. Люди, не предъявляющие жалоб или имеющие
четкую положи-тельную динамику, не нуждаются в таком обследовании.
Необходимо отметить, что в не-которых единичных случаях, даже при
наличии тяжелого заболевания, отмечается улуч-шение самочувствия и
исчезновение жалоб. Однако это улучшение недолгое и после пре-кращения
активационной терапии жалобы появляются вновь. Поэтому во второй анкете
необходимо примечание: при возобновлении жалоб следует обратиться к
врачу. Для диаг-ностического лечения применяются природные средства в
небольших дозах. При этом, когда возможно, используется сочетание
общего и местного действия (так, при заболеваниях желудка эффективен
кислородный коктейль с включением лечебных трав; при заболеваниях
дыхательной системы — ингаляции с растительным маслом,
прополисом, с травами в сочетании с «полным» дыханием,
дыхательной гимнастикой). При применении кислородных коктейлей или
ингаляций требование к новизне (по дозе), выполняется ав-томатически,
т. е. работает режим № 1.
Диагностическое лечение проводилось, во-первых, у людей в
донозологических со-стояниях, предъявляющих разнообразные жалобы, не
характерные для какого-либо забо-левания (124 человека). При
медицинском обследовании также не было поставлено ника-кого
определенного диагноза. Кроме того, диагностическое лечение было
проведено у 126 больных с заболеваниями желудка, имеющими диагноз
гастрит (гипо-, гипер- и норма-цидный) и 52 — с заболеваниями
дыхательной системы (чаще всего, бронхит).
Предварительно у всех исследовалось общее число лейкоцитов и
лейкоцитарная формула с подсчетом 200—300 клеток. У большинства
отмечался стресс разной тяжести (77%), у остальных —
адаптационные реакции тренировки и активации с элементами
на-пряженности (лейкоцитоз или лейкопения, моноцитоз, эозинофилия или
эозинопения, сдвиг влево, увеличение числа базофилов). Затем с учетом
характера заболевания или жа-лоб больным применялся тот или иной
биостимулятор по одному из программированных режимов. В легких случаях
давался режим № 1, в более тяжелых — режим № 2, при нали-чии
лейкопении — режим № 3. Если через
157
неделю после применения первого режима не было эффекта как
субъективного — четкого уменьшения жалоб, так и
объективного— выхода из стресса или уменьшения элементов
напряженности, пациентов на неделю переводили на второй режим. Если
также в течение недели не было эффекта при первоначальном использовании
второго режима, на неделю переводили на третий режим. Третий режим
обычно продолжали 1 — 1,5 недели. Послед-ние 2 недели почти все
больные (85%) проводили на первом режиме.
Через месяц более 80% людей с донозологическими состояниями отмечали
суще-ственное уменьшение или полное исчезновение жалоб. Анализы крови
укладывались в картину физиологических адаптационных реакций тренировки
и активации, самочувствие и работоспособность становились хорошими. Это
совпадает с данными литературы о том, что в «предболезни»
находится до 80% населения (Баевский Р. М., 1979;
Казначеев В. П., Баевский Р.
М., 1974).
Наблюдения в сроки от 10 месяцев до 1 года говорят об эффективности
получен-ных результатов. При периодически возникающих рецидивах
(вследствие психоэмоцио-нального напряжения, экстремальных ситуаций)
проводился повторный курс лечения также с хорошим результатом. У
небольшого процента людей, у которых диагностическое лечение не
вызывало четкого улучшения и исследование лейкоцитарной формулы также
свидетельствовало об отсутствии нормализации, были обнаружены
заболевания с выра-женными структурными изменениями: тяжелые
хронические гастриты, язвенная болезнь желудка и 12-перстной кишки,
хроническая пневмония. Хотя эти заболевания за период диагностического
лечения излечены не были, но все же отмечалось улучшение общего
состояния, уменьшение жалоб. Поэтому диагностическое лечение у этих
больных явилось как бы первым этапом лечения, подготавливающим организм
к специфической терапии и создающим для нее благоприятный
неспецифический фон.
Принцип диагностического лечения, простого и недорогого, проводимого
без от-рыва от производства, может быть полезен и для выявления групп
риска, и для лечения начальных стадий заболеваний. В комплексе с картой
здоровья, анкетами и другими про-стыми методами ранней диагностики этот
метод должен способствовать успеху дела дис-пансеризации.
Применение активационной терапии
на разных этапах лечения больных с опухолями
Для повышения эффективности лечения онкологических больных крайне важно
повысить неспецифическую резистентность организма, активность его
защитных систем. Существенно повышает резистентность организма
адаптационная реакция активации вы-соких уровней
158
реактивности (вызываемая малыми по абсолютной величине действующими
факторами). При химио- и лучевой терапии опухолей, нередко приводящих к
развитию лейкопении, более эффективна (для защиты от лейкопении)
реакция тренировки. Повышение рези-стентности организма необходимо на
всех этапах лечения онкологических больных, но на каждом этапе
используются разные методы вызова нужной в данное время адаптационной
реакции.
1. В период до начала специального лечения (операция,
химио-и лучевая терапия) следует вызывать развитие реакции
активации (а в случае исходной лейкопении — трени-ровки) с
помощью малых доз адреналина, мумие или
магнитного поля с малой вели-чиной
индукции. Величину действующего фактора подбирать по показателям
реакций — лейкоцитарной формуле (методические
рекомендации). Биостимуляторы растительного и животного
происхождения (элеутерококк, пантокрин и т.д..) менее эффективны, но
если подбор дозы по крови невозможен, полезно применять биостимуляторы,
подбирая дозу по режиму № 1 без
проведения частых анализов крови.
2. В послеоперационном периоде и при химио- и лучевой
терапии также наиболее эффективны адреналин и магнитное поле. В
послеоперационном периоде желательно раз-витие реакции активации путем
подбора дозы по лейкоцитарной формуле. При невозмож-ности этого
можно использовать биостимуляторы с
определением дозы по режиму № 1. При химио- и лучевой
терапии можно также подбирать величину МП и дозу адрена-лина по
лейкоцитарной формуле, вызывая при лейкопении реакцию тренировки, а при
нормализации общего количества лейкоцитов — реакцию
активации. Эффективно ис-пользование программированного режима №
3 (что также возможно без частых анализов крови). При этом могут
применяться и биостимуляторы (например, элеутерококк).
Для повышения эффективности лечения в случаях, когда отсутствует резко выра-женная лейкопения, может быть использован режим № 2.
3. В промежутках между специальным лечением и в процессе
реабилитации онко-логических больных необходимо повышать резистентность
организма, резко сниженную как опухолевой болезнью, так и побочным
действием специфической терапии.
В эти периоды можно применять 3 режима. Если еще есть лейкопения, то
начать нужно с режима № 3 — 1-1,5 месяца, а затем перейти на
режим № 1.
Если лейкопении нет, но есть стресс, то вначале дается режим № 2 — в течение 2 —3-х недель, а затем режим № 1.
Каждому больному после окончания стационарного лечения режимы выдаются на руки.
Соблюдение строгой этапности в лечении онкологических больных,
159
постоянное проведение терапии, повышающей резистентность организма,
активность его защитных систем должно существенно повысить
эффективность лечения и снизить коли-чество рецидивов и частоту
метастазирования.
Математические модели
Изучение периодической закономерности развития системы адаптационных
реак-ций и ареактивности, а также многочисленные экспериментальные
данные, полученные в процессе работы, позволили создать математические
модели. В основу первой, имитаци-онной модели динамики адаптационных
реакций, положена эвристическая модель моти-вации К. Лоренца (1966). В
отличие от модели Лоренца, имеющей концептуальный харак-тер, в данную
модель введена система дифференциальных уравнений (Файдыш Е. А., 1981).
При создании этой модели были использованы методы имитационного
моделирования, применяющиеся для решения экологических задач и изучения
иммунных процессов (Шеннон В., 1978; Марчук Г. И., 1980). В ходе
имитационного моделирования на ЭВМ получено, что при возрастании
интенсивности раздражителя происходит смена программ гомеостаза,
характеризующая динамику чередования основных адаптационных реакций,
показанную ранее в эксперименте. Данная модель позволяет разрабатывать
стра-тегии управления процессами гомеостаза, нацеленные на решение двух
задач: выведение из патологического состояния и поддержание хорошего
состояния здоровья с высокой ре-зистентностью (Гаркави Л. X., Квакина
Е. Б., Лупичев Л. Н., Файдыш Е. А., 1985). Вторая модель,
регрессионная, была создана на основе автоматизированной обработки
большого массива экспериментальных данных, характеризующих состояние
тимико-лимфатической и эндокринной систем, а также некоторых параметров
белой крови в соответствии с алго-ритмом корреляционно-регрессионного
анализа. По составу выходных данных, а также с помощью дискриминантной
функции были определены особенности формирования об-раза,
характеризующего реакцию активации. Данная модель позволяет определять
(про-гнозировать) эффективность воздействий по изменению доступных для
исследования единичных параметров за счет предсказуемости при этом (с
помощью модели) целого ря-да других параметров (Шихлярова А. И., 1985
— 1987). С помощью регрессионной моде-ли расчетным путем была
подтверждена надежность использования показателей процентного
содержания лимфоцитов в периферической крови в качестве информативного
сиг-нального теста развития комплекса изменений в организме после
воздействия.
Ю. В. Серегиным (1983, 1986) предложены метод и устройства для восстановления движений у детей с церебральным параличом,
160
а также при последствиях черепно-мозговой травмы и сосудистых
заболеваний головного мозга. Оптимальный лечебный эффект отмечается при
развитии реакции активации. Для определения нужных параметров
действующего фактора — электрического тока — Серегиным
разработана математическая модель развития неспецифических
адаптационных ре-акций организма при воздействии на головной мозг.
Использование количественных от-ношений между элементами белой крови
позволило описать реакции системой дифференциальных уравнений первого
порядка. При этом амплитуда тока рассчитывается с учетом, прежде всего,
числа лимфоцитов, а частота—с учетом других элементов
лейкоцитарной формулы. Серегин успешно применил разработанный им метод
на практике.
В заключение приводим некоторые из уже опубликованных работ с
применением системы адаптационных реакций в различных областях медицины.
Дальнейшее развитие и применение теории адаптационных реакций разных уровней реактивности в отдельных областях медицины.
В ряде работ показано, что тяжесть патологического процесса и
прогнозирование выздоровления находятся в тесной корреляционной связи с
типом адаптационной реак-ции. Так, был проведен ретроспективный анализ
историй болезни больных ишемической болезнью сердца (ИБС), находившихся
на стационарном лечении (Коломиевский Д. Л., 1982), который показал,
что ИБС протекает, чаще всего, на фоне стресса или напряжен-ных реакций
тренировки и активации, так что доля нормальных, физиологических
реак-ций составляет от 4 до 15%. При наиболее тяжелом течении болезни
регистрировалась ре-акция стресс, причем, чем тяжелее было состояние,
тем глубже был стресс: больше была выражена лимфопения. По мере
улучшения клинического состояния больных уменьша-лась доля стресса и
увеличивалось число физиологических реакций тренировки и актива-ции.
Представляет интерес проведенное Д. Л. Коломиевским сопоставление
прогности-ческой ценности показателей адаптационных реакций с другими
(в том числе, кардиоло-гическими) тестами. Чаще всего наблюдалась
корреляция между напряженными реакция-ми или стрессом — с одной
стороны, и, например, изменениями ЭКГ — с другой. Однако в тех
случаях, когда были расхождения (в единичных случаях у впоследствии
умерших людей), а именно: данные ЭКГ свидетельствовали о положительной
динамике, а данные анализа крови — о глубоком стрессе, показатель
адаптационной реакции оказывался бо-лее информативным. Автор заключает,
что показатели адаптационных реакций стресса, тренировки и активации
являются
161
высокоинформативными в комплексной оценке состояния больного ИБС,
инфарктом миокарда (ИМ) и динамики заболевания и могут быть
использованы в лечении и опреде-лении прогноза у больного ИБС и ИМ.
Корреляция тяжести заболевания ИМ с характером адаптационной реакции
сохранялась и в процессе физической реабилитации (Африканова Л. П.,
Бутов М. А., Денисова А. И., 1985).
Исследование адаптационных реакций у больных, перенесших ИМ, на
санаторном этапе реабилитации (Додис Л. И. и др., 1983) показало, что
больные, находящиеся в реак-ции активации, предъявляли меньше жалоб,
имели большую «толерантность к физической нагрузке (по данным
велоэргометрии и двигательных режимов). В этой группе было в 3 раза
больше ЭКГ с положительной динамикой, чем в «стрессорной»
группе, и почти в 2 раза меньше — с отрицательной динамикой и
отсутствием стабильности.
Исследование адаптационных реакций в более отдаленные сроки,
проведенное у больных мужчин, перенесших ИМ, показало, что развитие у
них реакции активации спо-собствует более быстрому восстановлению
обычного индивидуального сексуального по-ведения, что ведет к
психической и социальной реабилитации личности (Смирнов Г. В., 1983).
Характер адаптационной реакции оценивался в динамике у больных с
хронически-ми обструктивными бронхитами в стадии обострения,
находившихся на стационарном ле-чении. Более тяжелое течение
заболевания наблюдалось у лиц со стрессом, с напряжен-ными реакциями
тренировки и активации и с переактивацией. Наиболее тяжелые случаи
заболевания протекали на фоне стойкого высокоэтажного стресса с
глубокой лимфопени-ей, с эозинофилией. В этих случаях не только
отсутствовал лечебный эффект от проводи-мой (традиционной) терапии, но
процесс прогрессировал, присоединялись приступы уду-шья, развивалась
бронхиальная астма. Напротив, у больных с реакциями тренировки и
ак-тивации без элементов напряженности в лейкограмме лечебный эффект
был самым высо-ким: раньше восстанавливалась легочная вентиляция,
исчезали кашель и хрипы, уменьша-лась интоксикация, а в дальнейшем
наступала продолжительная ремиссия. У больных, выписавшихся в реакции
активации, не было обострений в течение года. Делается вывод о том, что
реакции тренировки, активации и стресса являются надежным
прогностическим тестом (Романов А. И., Стареева Л. В., Васильев В. В.,
Кайдаш А. Г. и др., 1983).
Анализ историй болезни лиц, страдающих ИБС, хронической пневмонией и
брон-хиальной астмой, часто и длительно находящих-ся на стационарном
лечении, выявил, что положительная динамика этих заболеваний
сопровождается переходом из стресса в реак-ции тренировки и активации,
а отрицательная — переходом в глубокий
162
стресс. Авторы делают заключение, что анализ типа адаптационной реакции
в клинике внутренних болезней является весьма перспективным методом
оценки состояния больно-го, повышает эффективность применяемого лечения
(Соколова Г. Б., Царькова М. Ю.. 1987).
При легочном туберкулезе не только определялся тип адаптационной
реакции, но и проводилась активационная терапия. Применялись адреналин,
левамизол, тималин в акти-вационной дозе (Рыбалко В. В., Браженко Н.
А., Хавинсон В. X., 1986). Нормализация гормонального статуса и
воспалительного потенциала организма при этом позволила от-казаться от
применения глюкокортикоидных гормонов.
Определение типа и характера адаптационной реакции у больных
туберкулезом да-ло возможность оценить характер и уровень
резистентности каждого больного и соответ-ственно проводить
патогенетическую активационную терапию. Изучена зависимость
адаптационных реакций от клинической формы туберкулеза, фазы процесса,
возраста и длительности туберкулостатистической терапии. На основе этих
исследований выявлено 4 типа неспецифической реактивности. Проведенная
оценка эффективности лечения ин-фильтративного и деструктивных форм
туберкулеза легких показала, что при целенаправ-ленном получении
физиологической, гармонической реакции активации (главным обра-зом,
спокойной активации) наблюдается наилучший эффект от проводимой
терапии: бы-строе рассасывание инфильтратов и рубцевание полостей
распада с формированием ми-нимальных посттуберкулезных изменений в
легких (Браженко Н. А., 1985).
Имеется сообщение об успешном применении активационной терапии в дермато-логии (Горголюк Ю. А., 1983).
В нейрохирургии активационная терапия применялась при нескольких видах
забо-леваний. О работе Ю. В. Серегина (лечение детского церебрального
паралича, черепно-мозговых травм и сосудистых заболеваний головного
мозга) говорилось ранее. Высокая эффективность была достигнута при
лечении оптохиазмального лептоменингита (Кикут Р. П., Лиепа М. Э.,
Кикуте С. Р. и др., 1981).
Ряд работ по изучению и коррекции адаптационных реакций был проведен в
хи-рургии. Известно, что хирургический (операционный) стресс может
рассматриваться как классическая модель стресса. Было показано, что
выраженное повышение резистентности к операционному стрессу достигается
при целенаправленном вызове реакции активации с учетом индивидуальной
чувствительности (Клецкин С. 3., 1983; Клецкин С. 3 и др., 1984).
В других работах оценка и коррекция состояния организма по характеру адаптационной реакции проводилась в предоперационном
163
периоде (Брюшков С. С, 1981, 1983). Традиционная предоперационная
подготовка выво-дила больных из острого стресса, но количество больных
в состоянии хронического стресса даже увеличивалось. Коррекция
адаптационных реакций (с помощью пирогенала) в предоперационном периоде
у больных язвенной болезнью, хроническим холециститом,
тромбооблитерирующими заболеваниями, а также онкологическими
заболеваниями I — II стадии оказала существенное влияние на
течение послеоперационного периода. Больные, которым наряду с
традиционной подготовкой вводили пирогенал, в послеоперационном периоде
быстрее выходили из стресса. Гнойные осложнения при традиционном
способе подготовки наблюдались в 12,5% случаев, при целенаправленном
выведении из стресса — только в 2,7%. Наиболее часто осложнения
развивались у больных, оперированных в со-стоянии острого и
хронического стресса. Влияние характера адаптационной реакции,
на-блюдающейся до операции, на течение послеоперационного периода
отмечена и при раке легкого (Клетке Г. Э., 1990).
Связь типа адаптационной реакции с клиническим течением заболевания и,
в том числе, с развитием поздних инфекционно-гнойных осложнений
обнаружена также при травматической болезни (Голубев Г. Ш. и др., 1983;
Голубев Г. Ш., Локшина Е. Г., 1985). При неосложненном течении травмы с
19-х суток стресс вообще не регистрируется, а пре-обладающей реакцией
является реакция активации. При осложненном течении травмати-ческой
болезни стресс является преобладающей реакцией на протяжении 30 суток с
момента
травмы.
Известная роль стресса в психический патологии и наряду с этим факты
нормали-зующего влияния на психику физиологических адаптационных
реакций послужили осно-ванием для разработки метода активационной
терапии психических заболеваний (Шевелев А. И., Невский М. П., Гаркави
Л. X., Уколова М. А., Квакина Е. Б., А. с. № 995692, 1982; Шевелев А.
И. и др., 1984). Исследования адаптационных реакций у больных
ши-зофренией, маниакально-депрессивным психозом, инволюционным
психозом, вторичны-ми энцефалитами показали, что у подавляющего
большинства этих больных преобладаю-щей реакцией является стресс. Для
вывода организма из стресса при всех этих заболева-ниях, т. е. с целью
вызова реакций тренировки и активации, были применены антидепрес-санты
(мелипрамин), гормоны (инсулин), адаптогены (элеутерококк), причем в
дозах в 7—20 раз меньших обычно применяемых. Так, доза
мелипрамина для получения реакции активации колебалась от 12,5 до 37,5
мг (терапевтическая доза —100 — 250 мг). Доза подбиралась
индивидуально каждому больному для получения и поддержания реакции
активации и тренировки по обратной связи с организмом. Клинические
наблюдения авто-ров свидетельствуют о том, что уже вскоре после
развития реакции активации
164
начинается улучшение психического состояния больных. При этом
симптоматические психотропные вещества назначались в дозах, в
3—10 раз меньших, чем при традиционном лечении. Эффективность
активационной терапии выражалась в сокращении времени пре-бывания
больного на койке. Выход из болезненного состояния (т. е. полное
исчезновение симптоматики, когда больные выписывались на работу, не
предъявляя никаких жалоб), наблюдался в 77,6% случаев, значительное
улучшение в 19,4 и улучшение — в 2,9% слу-чаев. В контроле, т. е.
без применения активационной терапии и с большими дозами пси-хотропных
препаратов, эти цифры составляли соответственно 34, 40 и 26%.
Возможность снижения высоких доз специальных медикаментов, далеко не
безразличных для организ-ма, является очень важным обстоятельством.
Е. И. Макеевой (1981) был разработан метод интенсивной активационной
психоте-рапии затяжных неврозов. Автор считает, что использование
теории адаптационных реак-ций дает возможность подойти с единых
теоретических позиций к выбору средств биоло-гического активирования,
осуществлять оперативный контроль функционального состоя-ния организма
и этим обеспечить реализацию активирования, соответствующую
возмож-ностям организма в данный момент времени.
Ю. М. Караш, Р. Б. Стрелков и А. Я. Чижов (1988) разработали новый
метод повы-шения неспецифической резистентности с помощью применения
прерывистой нормоба-рической гипоксии (использование в прерывистом
режиме газовой гипоксической смеси с пониженным количеством кислорода).
Используя новый метод, авторы переводили небла-гоприятные адаптационные
реакции (хронический стресс, переактивацию или напряжен-ные реакции
тренировки и активации) в физиологические реакции, дозируя газовую
смесь по числу циклов вдыхания и по концентрации в ней кислорода.
Переход в физиологиче-ские реакции сопровождался оздоровительным и
терапевтическим эффектом при ряде за-болеваний, повышением
адаптационных и резервных возможностей организма. Кроме этого,
изучалась роль адаптационных реакций в течении и прогнозе некоторых
заболева-ний.
Так, при хронических неспецифических рецидивирующих сальпингоофоритах
по-казана корреляционная зависимость между характером адаптационной
реакции, с одной стороны, и длительностью заболевания, частотой
обострений, тяжестью процесса, интен-сивностью болевого синдрома
— с другой (Караш Ю. М. и др., 1984; Караш Ю. М., Стрелков Р. Б.,
Чижов А. Я., 1988). Авторы полагают, что наличие комплекса изменений в
организме, характерного для хронического сальпингоофорита, разная
степень выраженно-сти функциональных нарушений, длительность течения,
частота обострений,
165
низкая эффективность комплексной терапии и склонность данного
контингента больных к воспалительным заболеваниям верхних дыхательных
путей — все это зависит от неблаго-приятных адаптационных реакций
организма. Последние отражают сниженную неспецифическую резистентность
и ослабление компенсаторно-приспособительных воз-можностей организма.
Применение больным с хроническим сальпингоофоритом, ранее длительно и
без-успешно лечившимся как амбулаторно, так и в стационаре,
гипоксической смеси в преры-вистом режиме под контролем адаптационных
реакций привело к значительному успеху. Более чем в половине случаев
наблюдалось полное выздоровление, в остальных терапев-тический эффект
оценивался как частичный. По мнению авторов, безуспешность ранее
проводившейся терапии была связана с тем, что она не включала в себя
метода контролируемого воздействия на общую резистентность организма.
Процент неблагоприятных реакций после примененного курса
нормобарической гипокситерапии снизился до 9,4±1,3 (вместо
60,8±3,8%. до лечения), а процент физиологических реакций
(спокойной и повышенной активации) увеличился до 90,6±4,1%. При
этом наблюдалась корреляция изме-нения характера реакции с эффектом
лечения. В группе с частичным клиническим эффек-том количество
неблагоприятных реакций резко уменьшилось: с 68,5±7,8% до
28,5±7,8%. После полного выздоровления у всех определялись
физиологические реакции спокойной или повышенной активации. При
достижении лечебного эффекта улучшались многие ла-бораторные
показатели, в том числе нормализовалась функциональная активность
лим-фоцитов за счет восстановления нормальной ферментативной активности
и перераспреде-ления субпопуляций Т-лимфоцитов в сторону увеличения
Т-хелперов. Отсюда заключе-ние авторов: «Возможность вызывать
целенаправленно физиологические адаптационные реакции способствует
формированию новых направлений в области профилактики, тера-пии и
реабилитации организма при различных заболеваниях, разработке методов
повыше-ния его устойчивости к неблагоприятным факторам внешней и
внутренней среды».
Сходные данные были получены и в наших работах (Гарка-ви Л. X и др.,
1982) при определений характера адаптационных реакций также у больных с
воспалительными про-цессами женской половой сферы в стадии обострения.
Во всех случаях до начала лечения отмечались нефизиологические реакции:
хронический стресс и напряженные (высокэтаж-ные) реакции тренировки и
активации. Исследование некоторых иммунологических пока-зателей
свидетельствовало о снижении активности показателей клеточного
иммунитета, особенно при хроническом стрессе. Проведение метода
активационной терапии (дозиро-ванные воздействия переменным магнитным
полем на биологически активные точки), в сочетании с
166
традиционным лечением неспецифического сальпингоофорита приводило к
сокращению курса лечения на 1/3 и уменьшению (в 5 — 10 раз)
количества потребляемых антибиоти-ков, аналгетиков и снотворных
(Захарова Н. П. и др., 1984).
Высокоэффективным оказалось использование (при таких же
гинекологических за-болеваниях) левамизола в дозах меньших, чем
терапевтические, под контролем вызова реакции активации. При этом
длительность лечения сокращалась также на 1/3 (Деев Э. А., Мелехина Л.
М., Мирович Д. Ю., А. с. № 1194419, 1985). Соответственно изменялся и
иммунный статус больных (Мелехина Л. М., 1988).
О применении нами активационной терапии в онкологии говорилось ранее. В
мо-нографии В. Г. Пашинского и К. В. Яременко «Проблемы
онкологической фармакотера-пии» (1983) отмечается необходимость
повышения резистентности организма путем раз-вития реакций активации
или тренировки на «профилактическом» этапе лечения
онколо-гических заболеваний у лиц, излеченных от злокачественных
новообразований, и в так называемых группах риска. Авторы используют
для этой цели биологически активные вещества природного происхождения
— адаптогены.
Защита организма от побочного действия полихимиотерапии в больших дозах
была получена с помощью магнитного поля при глиальных опухолях
головного мозга (Кикут Р. П. и др., 1981): всем больным удалось
провести массивную химиотерапию и уменьшить число переливаний крови в 5
раз при хорошем общем состоянии. Повторный компьютер-но-томографический
анализ показал прекращение роста или уменьшение объема опухоли. Средняя
продолжительность жизни была в 2,5 раза больше, чем при хирургическом
лече-нии. При этом, в отличие от больных контрольной группы, не только
не наблюдалось нервно-психических явлений угнетения, но напротив,
отмечалось приподнятое настрое-ние, а также улучшение сна и аппетита.
В другой работе специально исследовалось влияние характера
адаптационных ре-акций на течение послеоперационного периода у больных
раком легкого (Клетке Г. Э., Котляров Е. В., Бехтерева Е. И., 1984).
После хирургического лечения — радикальной операции
(пневмонэктомии или резекции легких) — было обнаружено, что
независимо от объема оперативного вмешательства, у всех больных после
операции развивался острый стресс, который на 5-е сутки начинал
переходить в реакцию тренировки и активации, хотя и напряженные
(особенно вначале). .Во всех случаях, когда после пульмонэктомии стресс
держался более 4-х суток, в последующем наблюдались бронхоплевральные
осложнения. Авторы считают, что наиболее благоприятное течение
послеоперационного периода отмечается при спокойной и повышенной
активации, нежелательными
167
являются как стресс, так и другая крайность — переактивация. До
операции у больных наблюдалось волнообразное изменение адаптационных
реакций, они были нестойкими, что, по мнению авторов, отражает борьбу
организма со стрессорным влиянием опухоли. Реакции тренировки и
активации были напряженными. Это находило отражение в содер-жании
изучаемых показателей. Количество Т-лимфоцитов, содержание альбумина,
альбу-мино-глобулиновый коэффициент были меньше, а содержание
фибриногена — больше у больных раком, чем у здоровых при любой
адаптационной реакции, в том числе и при ре-акциях тренировки и
активации, что говорит об их «высокоэтажном» характере.
Внутри же группы больных эти показатели были различными в зависимости
от типа реакции. Наименьшие значения Т-лимфоцитов, В-лимфоцитов и
альбумино-глобулинового коэф-фициента при наибольшем уровне фибриногена
отмечались при стрессе, противополож-ные соотношения — при
реакции активации, особенно повышенной (Клетке Г. Э., 1989).
Исследование связи между характером адаптационной реакции и иммунным
стату-сом при химио-лучевом лечении больных в далеко зашедших стадиях
рака гортани и по-лости рта показало, что даже в таких тяжелых случаях
состояние иммунитета существен-но улучшается при развитии реакций
тренировки и активации по сравнению со стрессом (Бирина Л. М. и
др., 1987).
Общее заключение, которое можно сделать на основании данных, приведенных в этом разделе, следующее:
1. Клиническое течение и прогноз при рассмотренных заболеваниях
коррелируют с адаптационными реакциями организма: наиболее тяжелое
течение и плохой прогноз, на-блюдаются при стрессе, переактивации и
высокоэтажных реакциях активации и тренировки, хороший —
при физиологических низкоэтажных реакциях без
признаков напря-женности,
особенно при реакции повышенной
активации.
2. Активационная терапия повышает эффективность традиционных
способов ле-чения, снижает повреждающее побочное действие специфических
препаратов, позволяет уменьшить их применение и является
неспецифической патогенетической терапией, так как в
основе любого заболевания,
помимо специфического компонента, лежит
на-рушение адаптационной деятельности организма и снижение
резистентности.
Однако основной задачей применения реакций тренировки и особенно
активации на разных уровнях реактивности .является управление
функциональным состоянием, ре-зистентностью организма. В большинстве же
тех работ, которые здесь приведены, воз-можности активационной терапии
далеко не исчерпаны. В некоторых работах адаптаци-онные реакции
использованы минимально. Тем не менее, даже такое применение давало
выраженный эффект.
168
К ТЕОРИИ АДАПТАЦИОННЫХ РЕАКЦИЙ
Попытаемся изложить свое общее представление о возникновении, характере
и значении различных общих неспецифических адаптационных реакций и их
роли в регуля-ции уровня неспецифической резистентности организма.
Одним из путей сохранения необходимого для жизни гомеостаза является
реагиро-вание — развитие общих адаптационных реакций. Наши
исследования показали, что раз-витие общих адаптационных реакций
подчинено количественно-качественному принципу: на различное количество
раздражителя организм отвечает качественно разными реакция-ми.
Изучаемые нами общие адаптационные реакции организма являются
неспецифиче-скими, т. е. возникают в ответ на действие различных по
качеству раздражителей опреде-ленной силы (дозы). Иными словами,
количество (мера, биологическая активность) явля-ется общим в действии
самых различных по качеству раздражителей и служит основой формирования
нескольких стандартных ответов организма. Качество раздражителя
накла-дывается на этот стандартный ответ как на основу.
Нами показано, что в ответ на действие различных по качеству
раздражителей фи-зиологических параметров развиваются неизвестные ранее
физиологические адаптацион-ные реакции: реакция тренировки — на
действие слабых раздражителей (Гаркави Л. X., 1969; Квакина Е. В.,
Уколова М. А., 1969) и реакция активации — на действие
раздражи-телей средней силы (Гаркави Л. X., 1968, 1969).
Каждая из этих реакций протекает стадийно и имеет свой определенный
комплекс изменений в организме, который определяет уровень и характер
резистентности.
Реакция тренировки имеет 3 стадии: стадию ориентировки, стадию перестройки, стадию тренированности.
В ЦНС преобладает охранительное торможение. В эндокринной системе
вначале умеренно повышается активность глюко- и минералокортикоидных
гормонов, а затем по-степенно увеличивается секреция
минералокортикоидов и нормализуется секреция глю-кокортикоидов на фоне
умеренно повышенной функциональной активности щитовидной железы и
половых желез. В первой стадии реакции тренировки повышается пассивная
ре-зистентность за счет снижения-чувствительности, а в третьей —
активная резистентность — за счет повышения активности защитных
систем организма: тимико-лимфатической, системы соединительной ткани.
Первая стадия реакции тренировки эффективна при острых воспалительных
про-цессах, для защиты от повреждающего действия облучения и
химиотерапии, особенно при выраженной лейкопении. Кроме того, нужно
отметить, что при реакции тренировки на-блюдается мягкий, но четкий
антикоагуляционный эффект. Степень повышения
169
резистентности в этой стадии недостаточна для получения
противоопухолевого действия. В третьей стадии степень повышения
резистентности больше, повышается и противоопу-холевая резистентность.
Однако практическое использование этой стадии для лечения по-ка
затруднено, так как для достижения ее необходима длительная
систематическая трени-ровка.
Реакция активации, имеет 2 стадии: стадию первичной активации и стадию
стойкой активации. В ЦНС преобладает умеренное, физиологическое
возбуждение. В эндокринной системе наблюдается увеличение секреции
минералокортикоидов при нормальной секреции глюкокортикоидов и
повышении функциональной активности щитовидной железы и половых желез.
Повышение активности желез внутренней секреции выражено больше, чем при
реакции тренировки, но не носит характера патологической гиперфункции.
В обеих стадиях реакции активации повышается активная резистентность к
повреждающим агентам различной природы. С помощью развития реакции
активации удается получить торможение роста вплоть до полного
рассасывания уже развившихся экспериментальных опухолей как перевивных,
так и первично индуцированных. Необходимо отметить, что
противоопухолевого эффекта удается добиться только при систематическом,
каждоднев-ном повторении активационных воздействий, т. е. при
поддерживании стойкой активации организма в течение длительного
времени, вплоть до полного рассасывания опухоли.
Реакция активации подразделяется на спокойную активацию (РСА) и
повышенную акти-вацию (РПА). РПА вызывается раздражителями, несколько
большими по абсолютной ве-личине, чем РСА. Эти реакции удалось выделить
благодаря большему повышению актив-ности тимико-лимфатической системы,
большему относительному лимфоцитозу и более высокому уровню
резистентности при РПА, а также по разному влиянию РПА и РСА на систему
свертывания крови: нормализации при РСА и сдвигу в сторону
гипокоагуляции — при РПА, влиянию на артериальное давление,
уровень глюкокортикоидных гормонов, энергетику и др. Кроме того, РПА
отличается от РСА значительно большей стойкостью.
Особенно повышается резистентность в РПА. С помощью РПА в эксперименте
удалось получить рассасывание опухолей даже больших сроков развития,
что считалось ранее принципиально невозможным. Однако РПА опасна тем,
что при наличии тяжелого патологического процесса даже небольшая
передозировка приводит к развитию стресса.
Реакция активации эффективна при хронических воспалительных процессах,
для защиты от повреждающего действия различных токсических препаратов,
чрезмерных на-грузок. Реакция активации способствует нормализации как
повышенного, так и снижен-ного артериального давления.
170
Реакция тренировки и реакция активации — это те адаптационные
реакции, кото-рые встречаются в течение нормальной жизни организма.
Считая стресс единственной неспецифической адаптационной реакцией,
многие исследователи не определяли параметры реакции, доказывающие
развитие стресса. Г. Се-лье четко говорит о наличии при всех видах
стресса реакции тревоги со всем симптомо-комплексом, позволившим ему в
свое время (в 1936 г.) открыть эту адаптационную реак-цию. Отсутствие
реакции тревоги, замена свойственного ей симптомокомплекса на дру-гой,
с антистрессорным характером изменений отличает адаптационные реакции
трени-ровки и особенно активации. Открытие Г. Селье реакции стресс,
являющейся неспецифи-ческой основой патологических процессов —
«синдромом болезни вообще» (Селье Г., 1973), способствовало
пониманию общности в течение различных патологических про-цессов,
вооружило медицину знанием теории, помогающей не только вскрыть
патогенез, но и обосновать терапию целого ряда болезней. Однако
дальнейшее развитие учения об адаптационных реакциях организма привело
к необходимости четко выделить место стресса среди других адаптационных
реакций (Гаркави Л. X., 1968, 1969; Гаркави Л. X., Квакина Е. Б.,
Уколова М. А., 1979).
Считают, что в настоящее время на основе стресса развиваются около
десяти тысяч заболеваний и более ста тысяч болезненных симптомов
(Тигранян Р. А., 1988). Существу-ет мнение, что все нарушения в
организме имеют психосоматически-соматопсихическую природу (Temoshok
L., 1983),— с этим и может быть связан столь обширный спектр
по-ражений при стрессе.
Наши исследования подтвердили, что при опухолевой болезни развивается
хрони-ческий стресс. С этой точки зрения мероприятия по борьбе с
заболеваниями должны включать в себя борьбу со стрессом.
Какие же существуют методы борьбы со стрессом? В иностранной литературе
ши-роко применяется термин coping, введенный Лазарусом (Lazarus
R. S., 1966), — преодо-ление (стресса). Рост технизации,
ускоряющийся темп жизни, увеличение психоэмоцио-нального напряжения на
фоне гиподинамии, избыточность информационных нагрузок, появление новых
факторов, в том числе токсических, к которым организм не имеет
эво-люционно выработанных мер защиты — все это является причиной
роста stress of life, стресса жизни. Особенно большое значение в
возрастании «стресса жизни» в настоящее время придают
эмоциональному стрессу. Предлагаются различные способы копинга —
от изменения отношения к окружающему, т. е. собственной реакции на
стрессирующий фак-тор, до использования транквилизаторов, алкоголя,
наркотиков. Недостатки этих методов
171
очевидны. Транквилизаторам посвящена
обширная литература. Об их роли при стрес-се можно сказать, что
эпизодически или кратковременно она может быть положительной, но
рекомендовать транквилизаторы в качестве постоянной меры борьбы со
стрессом опасно — нужно иметь в виду и интоксикацию, и
привыкание, и потерю правильной оценки ситуации (Сох Т., 1978).
Опасность использования наркотиков не нуждается в специальном пояснении.
Что касается личностного восприятия ситуации, то при стойком стрессе
его изме-нить трудно, если не невозможно. В работах Л. Тимошек
подчеркивается, что для разви-тия стрессовых изменений большее
значение, чем свойства стрессора, имеет текущая оценка ситуации,
зависящая от целого ряда обстоятельств, связанных с особенностями
ор-ганизма данного человека (Temoshok L., 1983). По нашему мнению,
повлиять на отноше-ние к окружающему и этим содействовать копингу можно
путем перевода стресса в дру-гую, также адаптационную, также
неспецифическую реакцию, но, в отличие от стресса, физиологическую
— активации (лучше всего повышенной) или тренировки, что
сопрово-ждается изменениями психики, свойственными этим реакциям,
отличающимися меньшей тревожностью и большей оптимистичностью, чем
стресс.
Вместе с тем распространены представления о повышении резистентности с
помо-щью стресса и об. использовании стресса для закаливания,
спортивных тренировок.
Целесообразно ли повышать резистентность с помощью стресса?
Как уже говорилось, за счет стресса повышается, главным образом,
пассивная ре-зистентность, благодаря развитию запредельного торможения
и снижению чувствитель-ности, причем происходит это ценой больших
повреждений и энергетических трат. Мы думаем, что активная
резистентность при стрессе повышается, если повторные стрессор-ные
воздействия ввиду снижения чувствительности вызывают развитие реакции
актива-ции, т. е. не за счет стресса. Однако даже если с помощью
стресса можно повысить актив-ную резистентность, то и в этом случае
эффект лучше получить с помощью других реак-ций — без явлений
повреждения и с меньшими энерготратами. Поэтому мы считаем, что
использовать стресс — реакцию, лежащую в основе патологических
процессов,— с ле-чебной целью можно лишь в особых случаях. Широко
используется введение глюкокор-тикоидных гормонов в дозах, оказывающих
провоспалительное действие, подобно первой фазе реакции стресс. Однако,
учитывая отрицательное влияние этой стадии стресса на за-щитные системы
организма — угнетение тимико-лимфатического аппарата, системы
со-единительной ткани, иммунодепрессию — глюкокортикоиды должны
применяться крайне осторожно. Что касается стремления использовать
стресс при спортивных тренировках или закаливании, то это противоречит
простой логике. По мнению целого ряда исследова-телей, с
которыми мы полностью согласны, стресс
172
самой различной природы и так встречается в жизни слишком часто. Стресс
в трениро-вочном процессе, как и в процессе закаливания, может
исчерпать резервы организма и создать неблагоприятный
психоэмоциональный фон, привести к срыву и т. п. Показано, что при
закаливании оптимальной является реакция активации (Бутов М. А.,
Ногаллер А. М., 1986).
Адаптационные реакции формируются в ЦНС. По нашему мнению, способность
ЦНС реагировать как единое целое, вызывая развитие в организме общих
неспецифиче-ских реакций, связана с измененной, усложненной в процессе
эволюции, но диффузной по характеру реагирования частью ЦНС. Возможно,
ретикулярная формация является мор-фологическим субстратом,
объединяющим различные отделы мозга, принимающих уча-стие в
формировании адаптационных реакций организма. Исследования показали,
что важнейшая роль в формировании адаптационных реакций принадлежит
гипоталамусу — регулятору внутренней среды организма.
Специфическое и неспецифическое тесно пере-плелось в мозгу, и ни одна
специфическая функция не может реализоваться без опреде-ленного
неспецифического фона. Характер этого фона, по нашему мнению,
определяется силой (дозой) раздражителя. Это биологически
целесообразно, так как позволяет отвечать на воздействие адекватно, без
лишних трат.
При действии сильных, чрезвычайных раздражителей (реакция стресс) в ЦНС
раз-вивается резкое возбуждение, сменяющееся запредельным торможением
— крайней ме-рой защиты. Биологическая целесообразность этого
— в снижении возбудимости, реак-тивности, так как адекватный
чрезмерной силе раздражения ответ мог бы погубить орга-низм. Затем,
вследствие снижения реактивности, сильные воздействия уже не
восприни-маются как сильные, развивается стадия резистентности. Этим и
объясняется, что измене-ния, развивающиеся в стадии резистентности
стресса, как это ни парадоксально, связаны не со стрессом. Снижение
возбудимости при развитии запредельного торможения приво-дит к тому,
что сильные раздражители (в случае повторения действия стрессора) уже
не являются для организма сильными и вызывают развитие не стресса, а
реакции активации или даже реакции тренировки. Если же действие
стрессора не повторяется и на организм падают обычные раздражители
физиологических параметров, развивается чаще реакция тренировки, но
возможно развитие и реакции активации. По-видимому, с неоднородно-стью
стадии резистентности при стрессе связано отсутствие четкой
характеристики эн-докринной и тимико-лимфатической системы в этой
стадии по сравнению с реакцией тре-воги. Поскольку при стрессе стадия
резистентности развивается после реакции тревоги — стадии, для
которой наряду с элементами защиты характерно наличие повреждения
за-щитных систем и больших энергетических
173
трат, можно согласиться с Г. Селье, что резистентность при стрессе
оплачивается «доро-гой ценой» (Селье Г., 1961). Если
действие стрессора систематически повторяется или ра-зовый стрессор был
чрезвычайно сильным, стадия резистентности переходит в стадию
ис-тощения, которая может привести к гибели.
При действии слабых, пороговых (для общих реакций) раздражений
развивается реакция тренировки. В ЦНС при этом преобладает
охранительное торможение. Биологи-ческая целесообразность этого —
в снижении возбудимости, реактивности по отношению к слабому
раздражителю, на который наиболее целесообразно не отвечать.
При действии раздражений средней силы происходит развитие
«реакции актива-ции». В мозгу преобладает умеренное
возбуждение. По-видимому, раздражение средней силы является оптимальным
для возбуждения защитной деятельности организма. На такое раздражение
наиболее целесообразно реагировать путем первичной активации защитных
систем организма.
Таким образом, в формировании каждой адаптационной реакции участвует и
ЦНС, в особенности регулятор внутренней среды — гипоталамическая
область мозга и эндок-ринная система.
Мысль А. Д. Сперанского (1935) о том, что нервная система организует
патологи-ческий процесс, получает новое освещение с позиции теории
адаптационных реакций ор-ганизма. Все адаптационные реакции формируются
в ЦНС, в частности в гипоталамусе. В ЦНС формируется и реакция стресс,
являющаяся неспецифической основой патологиче-ского процесса, т. е. с
этой точки зрения, ЦНС организует патологический процесс. Там же
формируются реакции тренировки и активации, являющиеся неспецифической
осно-вой нормы и повышающие неспецифическую резистентность, т. е.,
иными словами, нерв-ная система организует и защиту от патологических
процессов.
Часто задают вопрос, чем же отличаются адаптационные реакции тренировки
и ак-тивации от реакции стресс, если в их развитии также участвует
гипоталамо-гипофизарно-надпочечниковая система? Как уже говорилось,
общие неспецифические адаптационные реакции — это такие реакции,
с Помощью которых организм, реагируя, сохраняет необ-ходимое для жизни
относительное постоянство внутренней среды. В мозгу самое
непо-средственное отношение к регуляции внутренней среды имеет
гипоталамус, осуществ-ляющий эту регуляцию через вегетативный отдел
нервной системы и эндокринную систе-му. Других систем для регуляции
внутренней среды в организме нет. То обстоятельство, что эта система
может меняться по-разному при действии раздражителей различной силы, т.
е. формировать развитие различных адаптационных реакций, позволяет
организму гиб-ко, биологически целесообразно приспосабливаться к
многообразным воздействиям
174
внешней и внутренней среды.
По нашему же мнению, адаптационные реакции организма подчиняются
принципу работы функциональных систем, которые, согласно П. К. Анохину
(1975), складываются из динамически мобилизуемых структур в масштабе
целого организма. Поскольку эти ре-акции сложились в процессе эволюции,
оптимальный вариант реакции, позволяющий со-хранить гомеостаз с
минимальными затратами, уже заранее известен, запрограммирован в
акцепторе результатов действия. В процессе афферентного синтеза
доминирующей мотивацией является сохранение гомеостаза оптимальным
путем. Память в этом случае связа-на, прежде всего, не с индивидуальным
опытом, а с опытом поколений. Обстановочная афферентация складывается
на основе синтеза ситуации, имеющейся в данный момент внутри организма
и в окружающей его среде. Пусковая афферентация обусловлена
раз-дражителем, вызывающим развитие той или иной адаптационной реакции.
При развитии адаптационной реакции так же, как и при формировании любой
функциональной системы, компоненты тех или иных анатомических
образований вовлекаются по принципу взаимо-СОдействия, направленного на
получение оптимального полезного результата.
Как уже говорилось, комплекс изменений, присущих каждой адаптационной
реак-ции, определяет характер и уровень резистентности, а также
преобладание провоспали-тельного или противовоспалительного потенциала
в организме. Что подразумевается под характером резистентности?
Резистентность может быть активной, основанной на различ-ной активности
защитных систем организма, или пассивной, основанной на различной
чувствительности, реактивности ЦНС. Иными словами, более устойчив тот
организм, ко-торый или лучше сопротивляется, или менее чувствителен.
Уровень резистентности опре-деляется степенью защитного действия по
отношению к различным повреждающим фак-торам. О преобладании
провоспалительного или противовоспалительного потенциала можно судить
по влиянию адаптационных реакций на течение острого и хронического
воспаления.
При реакции активации происходит умеренное повышение провоспалительного
потенциала организма. Поэтому развитие реакции активации показано при
хронических вялотекущих воспалительных процессах.
Необходимо подчеркнуть, что чередование стадий реакций тренировки и
активации про-исходит при систематическом повторении тренировочных или
активационных воздейст-вий. В то же время даже разовое применение
стрессора может привести к развитию всех трех стадий стресса. Однако
стадия истощения при разовом воздействии стрессора, как известно,
развивается только если стрессор очень сильный. В других случаях
отмечается
175
развитие реакции тревоги и стадии резистентности, т. е. после реакции
тревоги происходит возвращение к одному из вариантов нормы. Как уже
говорилось, стадию резистент-ности мы относим не к стрессу, а к
развитию вследствие запредельного торможения (сни-жения возбудимости)
либо реакции тренировки, либо активации. Иными словами, мы считаем, что
все эти реакции имеют ритм, близкий к околосуточному. При разовом
при-менении тренировочных или активационных воздействий развивается
стадия ориентиров-ки или стадия первичной активации и затем стадия
возвращения к состоянию, свойственному данному организму.
Закономерное развитие всех трех стадий стресса, включая стадию
истощения, на-блюдается также при систематическом повторении
стрессорных воздействий.
Обнаружение периодической (циклической) закономерности развития
адаптацион-ных реакций и состояний ареактивности существенно расширяет
и углубляет теоретиче-ские представления об адаптационной деятельности
организма.
К ТЕОРИИ МНОГОУРОВНЕВОЙ РЕГУЛЯЦИИ
ГОМЕОСТАЗА ОРГАНИЗМА С ПОМОЩЬЮ
ПЕРИОДИЧЕСКОЙ СИСТЕМЫ НЕСПЕЦИФИЧЕСКИХ
АДАПТАЦИОННЫХ РЕАКЦИЙ И АРЕАКТИВНОСТИ
В чем суть развиваемой нами теории адаптации, регуляции гомеостаза организма?
В процессе эволюции сложились две системы приспособления живого
организма, поддержания и регуляции необходимого для жизни гомеостаза:
система неспецифических адаптационных реакций и система ареактивности
(отсутствия реакции). Эти системы от-ражают два принципа
приспособления: принцип реагирования и принцип отсутствия ре-акции
(Гаркави Л. X., Квакина Е. Б., 1988). Деятельность обеих систем
подчинена перио-дической количественно-качественной закономерности
(Гаркави Л. X., Квакина Е. В., 1975— 1988). По мере возрастания
(уменьшения) величины любого действующего факто-ра на определенную
величину (коэффициент реакции) в организме периодически повто-ряются
разные по качеству тетрады основных адаптационных реакций: реакция
трениров-ки (Гаркави Л. X., 1969; Квакина Е. В., Уколова М. А., 1969),
реакция активации (Гаркави Л. X., 1968, 4969), подразделяемая затем на
реакцию спокойной активации и реакцию по-вышенной активации, и стресс
Г. Селье (1936).
176
Система ареактивности также подчинена количественной закономерности, но
по другой шкале отсчета величины действующего фактора. В реализации
этой системы осо-бое значение имеет повторность воздействий.
Вначале была найдена более простая зависимость развития адаптационных
реакций от интенсивности (силы, дозы) действующего фактора: реакция
тренировки — на малые, активации — на средние, стресс
— на большие по абсолютной величине раздражители.
В процессе изучения механизмов развития и способов вызова в организме
различных адаптационных реакций выяснилось, что
количественно-качественная закономерность не ограничивается однократной
сменой адаптационных реакций тренировки, активации и стресса по мере
увеличения величины воздействия. Определился циклический,
периоди-ческий характер развития неспецифических адаптационных реакций
организма: по мере увеличения (или уменьшения) интенсивности (силы,
дозы) различных по качеству действующих факторов отмечалось
периодическое повторение основных тетрад адаптацион-ных реакций
(тренировка, спокойная активация, повышенная активация, стресс),
разви-вающихся на разных уровнях реактивности (Гаркави Л. X., Квакина
Е. В., 1975 — 1985). Представления об уровнях реактивности
просты: если организм способен реагировать на очень малые по абсолютной
величине действующие факторы, это означает, что реактив-ность организма
высокая, если же организм способен реагировать на большие по
абсо-лютной величине факторы, реактивность организма низкая. На самом
деле, как известно, живой организм может реагировать на разные по
абсолютной величине действующие факторы, от очень малых до больших, т.
е. реагировать на разных уровнях реактивности. На каждом уровне
реактивности слабые (для данного уровня) действующие факторы вы-зывают
развитие реакции тренировки, средние — реакции спокойной и
повышенной акти-вации, сильные — стресс.
Периодический характер обнаруженной зависимости развития основных
адаптаци-онных реакций от силы (дозы), по-видимому, является одним из
проявлений общей пе-риодической закономерности, существующей на разных
уровнях организации материи, начиная с химических элементов,
подчиняющихся периодическому закону Д. И. Менде-леева. В основе такой
закономерности лежат дискретные количественные изменения раз-личных
свойств. Например, для отдельных атомов и химических элементов —
это перио-дическое повторение сходных типов электронных конфигураций и
химических свойств по мере возрастания атомного номера. На примере
адаптационных реакций — это периоди-ческое повторение сходных
(одноименных) реакций но мере возрастания количества дей-ствующего
фактора. Весь «набор» известных типов адаптационных реакций
можно срав-нить с периодами, а одноименные реакции разных
177
уровней реактивности — с группами Периодической системы элементов Д. И. Менде-леева.
Обнаруженная закономерность имеет свои количественные характеристики.
Опре-делился коэффициент реакций (К) — величина (индивидуальная),
на которую нужно из-менить величину (дозу, силу) действующего фактора,
чтобы перейти в «соседнюю» реак-цию. К оказался весьма
небольшой величиной — от 1,1 у больных и пожилых до 1,3 у
здоровых и молодых. Чаще всего встречается К, равный 1,2, а 1,3 —
крайне редко. Такая закономерность подразумевает наличие дискретности
— многих дискретных состояний, сменяющих друг друга при очень
небольшой величине изменения действующих факторов (на
10—20—30%). Это видно на примере разнонаправленных
изменений различных пока-зателей по мере перехода из одной реакции в
другую (процентное содержание лимфоци-тов, содержание глюкокортикоидов,
минералокортикоидов, гормонов щитовидной и по-ловых желез и т. п.) и с
одного уровня реактивности на другой. Известно высказывание Ф. Энгельса
о том, что в природе нет скачков, потому что она целиком состоит из
скачков.
Никоим образом не отрицая роли и наличия специфического компонента,
связан-ного с качеством действующего фактора, одновременно с
неспецифическим компонентом, связанным с количеством этого же фактора,
мы придаем количественному параметру ос-новное значение в формировании
нескольких стандартных адаптационных реакций, раз-вившихся в процессе
эволюции. Именно количество (мера биологической активности для живого)
есть то общее, что объединяет действие различных по качеству факторов и
созда-ет реальную основу для формирования общих ответов. Кроме того,
нужно помнить, что живое реагирует — и весьма тонко —
прежде всего на изменение. Изменения по количе-ственному параметру
являются общими для факторов, самых различных по качеству, и также
могут явиться основой развития общих для разных факторов изменений.
Ганс Селье, обладавший большой интуицией, предчувствовал наличие,
помимо стресса, дистресса, который он характеризовал как
«скорость изнашивания организма» (Селье Г., 1987), наличие
иного функционального состояния, более благоприятного —
эу-стресса, но, как уже говорилось, отсутствие объективных
количественных критериев эу-стресса не дало возможности разграничить
эти состояния. Очевидно, имея в виду положи-тельные черты эустресса,
Селье высказывает мысль о том, что жизнь без стресса есть смерть.
Безусловно, чтобы не впасть в большую ошибку, это положение можно
трактовать только как то, что полноценная жизнь невозможна без
необходимой степени активации.
В западной литературе появилось в последнее время понятие
«активация», под ко-торой подразумевается любое приращение
активности в эндокринной системе (Ursin H.,
178
1978; Ursin H et al., 1980). Свое представление об активации Урсин
связывает с известным, можно сказать, классическим
электрофизиологическим феноменом: десинхронизацией ЭЭГ, регистрируемой
при активном бодрствовании — «реакцией активации»
коры голов-ного мозга, но G. Моruzzi и Н. Magoun (1949). Рассмотрение
гормональной и поведенческой активности с учетом изменений активности
ЦНС является, безусловно, плодотвор-ным, так как связывает
функциональное состояние мозга с функциональным состоянием организма.
Авторы имеют также в виду данные Мэйсона о влиянии изменений в
психо-эмоциональной сфере на эндокринную систему (Mason J. W., 1971).
Изучались гормо-нальные и биохимические показатели у лиц, обучавшихся в
военной школе парашютистов (Levin S., Weinberg J., Ursin H., 1978;
Levin S., 1978; Ellertsen В., Jonsen Т., Ursin H., 1978). В процессе
исследований производилось сравнение этих показателей после первых
прыж-ков, когда преобладающим было чувство страха, и на этапах
дальнейшего обучения, когда страх у обучающихся шел на убыль (что,
кстати, сопровождалось более успешным вы-полнением прыжков). Те
изменения гормональной активности («активация» по
термино-логии авторов), которые были описаны в первый период —
повышение уровня кортизола, адреналина, норадреналина, гормона роста и
глюкозы наряду со снижением уровня тесто-стерона,— фактически
укладываются в картину стресса. У лиц, которые в процессе обу-чения
отсеялись (не смогли преодолеть чувства страха), такая картина
сохранялась и в по-следующем. Сравнение показателей деятельности
эндокринных желез с поведенческими реакциями позволило авторам сделать
вывод о том, что первоначальная «активация» уменьшалась,
когда человек справлялся с ситуацией. При этом повышенная вначале
гор-мональная активность снижалась, сниженная — повышалась. Это
расценивалось как то, что по мере успешного обучения достигался копинг.
Следовательно, под копингом авторы понимают уменьшение первоначальной
«активации» несмотря на то, что действие факто-ров,
заключающих в себе опасность или даже угрозу для жизни, не
прекращается. Эти весьма интересные данные тем не менее свидетельствуют
о том, как трудно оценить функциональное состояние организма, если не
иметь в виду количественного подхода. Урсин сам отмечает, что если
активация — «только активация», то это полезно, а
если она оказывается стрессом (!), то это чревато патологией.
Фактически вначале (первые прыж-ки) речь шла о стрессе (так же, как и в
дальнейшем у лиц, оказавшихся неспособными к этому роду деятельности),
а затем могли развиваться разные состояния: от стрессорной
ареактивности — отсутствия реакции на продолжающееся воздействие,
до развития реак-ции активации и, скорее всего, повышенной, поскольку
априори можно полагать, что у
179
молодых людей, успешно справлявшихся с описанным заданием, чувство
страха может сменяться сильными положительными эмоциями. Кстати
сказать, успешное выполнение задач такого типа, как парашютные прыжки,
должно обязательно сопровождаться именно повышенной активацией. По
нашему мнению, в процессе обучения происходил копинг стресса именно с
помощью реакции повышенной активации (в данном случае даже невы-соких
уровней реактивности).
Исследования, в которых при оценке реакции на раздражитель принимается
во внимание, а тем более строго учитывается его количественная сторона,
чрезвычайно ред-ки. Одним из исключений является работа М. Вигаша,
который даже повреждающие фак-торы— травму, эндотоксины, острые
кровотечения — рассматривает в качестве стрессор-ных только в тех
случаях, когда они достигают определенной интенсивности. Вигаш
счи-тает, что стресс развивается, если действующий фактор является либо
количественно чрезмерным, либо качественно стрессогенным и представляет
опасность для целостного организма — реальную или символическую
(Vigas M., 1980, 1984). Действительно, такие критерии могут быть
приняты для характеристики
«высокоэтажного» стресса —
стресса Селье.
Значительно более часто в работах по изучению стресса любые
воздействия, неза-висимо от их интенсивности, априори считаются
стрессорными (иммобилизация — не очень длительная, болевые
раздражения — не очень сильные и другие факторы, которые могут
вызывать самые разные реакции в зависимости от их интенсивности,
длительности действия, исходного состояния, индивидуальной
чувствительности). Такой подход связан, по-видимому, с привычным
представлением о существовании лишь одной неспецифиче-ской
адаптационной реакции — стресса. Однако у некоторых
исследователей возникает недоумение: каким образом даже минимальное
воздействие на организм, которое никак не может быть стрессорным,
способно запустить нейроэндокринные реакции, повышающие резистентность
организма даже в экстремальных условиях? (Тигранян Р. А., 1988). По
ви-димому, автор столкнулся с «тренировочными» или, что
более вероятно, «активационны-ми» воздействиями, причем
малыми по абсолютной величина.
Иными словами, отсутствие четкого количественного подхода к
оценке влияния действующих факторов затрудняет возможность градации
функциональных состояний организма, изучения и целенаправленного
применения неспецифических адаптационных реакций, каждая из которых
характеризуется присущими ей чертами и в соматической, и психической
сфере и играет определенную роль в адаптации организма,
как кратко-, так и долговременной.
Чрезвычайно важным для понимания механизмов долговременной адаптации
явля-ется положение Ф. 3. Меерсона, получившее широкую
известность, о формировании
180
«системного структурного следа» — комплекса
структурных изменений, увеличивающих мощность функциональной системы,
доминирующей при действии данного фактора. Структурный след, согласно
концепции Ф. 3. Меерсона (1981, 1986), составляет основу долговременной
фенотипической адаптации. Образование структурного следа происходит
вследствие активирующего влияния повышенной функции через генетический
аппарат клетки на процессы синтеза нуклеиновых кислот и белков. По
мнению Ф. 3. Меерсона, ведущим звеном в этом механизме является
стресс (Меерсон Ф. 3., 1986).
На основании наших исследований можно в развитие данной концепции
добавить, что структурный след формируется в результате развития не
только стресса, но и других реакций, причем при каждой из них —
свой. Преобладание процессов анаболизма при по-вышенной активации
должно способствовать быстрому формированию структурного сле-да,
необходимого для эффективной адаптации.
В работах Ф. 3. Меерсона убедительно показано, что предварительная
адаптация к воздействиям, меньшим по величине, чем стрессорные, к
«коротким стрессорным воздей-ствиям», по терминологии
автора, предотвращает тяжелые стрессорные повреждения миокарда, в том
числе, ишемические. Говорится также о том, что такого рода эффекты
реализуются лишь при «рациональном дозировании» и
«адекватном подборе» физических нагрузок (иначе последствия
могут быть и отрицательными). В таком адаптированном ор-ганизме
увеличивается активность «стресс-лимитирующих» систем
(Меерсон Ф. 3., Пшенникова М. Г., 1988). На основании наших многолетних
исследований можно пред-полагать, что в особенно успешных случаях
профилактики стрессорных поражений пред-варяющее воздействие должно
быть, по нашей терминологии, активационным. В безус-пешных же случаях
это может быть и стресс.
Что касается стресс-лимитирующих систем (Меерсон Ф. 3., 1986), то мы
думаем, что во многом благодаря существованию таких систем и возможно
развитие других реак-ций, помимо стресса,— реакций
антистрессорной защиты организма. Но в таком случае активность этих
систем также нуждается в количественной градации: избыточная
актив-ность может привести к развитию неблагоприятной реакции —
если не стресса, то переак-тивации. Таким образом, концепции Ф. 3.
Меерсона о системном структурном следе и стресс-лимитирующих системах
находятся в тесной взаимосвязи с концепцией о
количе-ственно-качественной закономерности развития адаптационных
реакций. По нашему мне-нию, идентификация характера адаптационной
реакции при применяемых Меерсоном воздействиях (хотя бы по самому
простому сигнальному показателю реакций — специ-альным образом
подсчитанной лейкограмме) дала бы новые представления о состоянии
стресс-лимитирующих систем при разных реакциях и состояниях
ареактивности. Это, в
181
свою очередь, позволило бы более дифференцированно подойти к подбору
величины дей-ствующего фактора, наиболее адекватной для защиты от
стресса.
Сравнение одноименных реакций на разных уровнях реактивности показало
нали-чие черт сходства, позволяющих сохранить название реакции, и черт
различия. Так, оста-лось общим соотношение функциональной активности
эндокринных желез.
При стрессе характерно увеличение глюкокортикоидов и снижение
минералокор-тикоидов, снижение функциональной активности щитовидной и
половых желез и угнете-ние функциональной активности
тимико-лимфатической системы. Однако выраженность этих изменений
невелика при мягком стрессе (стрессе высоких уровней реактивности) и
значительна при тяжелом стрессе низких уровней.
При развитии реакции активации отмечается умеренное увеличение секреции
ми-нералокортикоидов при уровне глюкокортикоидов на нижней границе
нормы (спокойная активация) или в пределах верхней половины зоны нормы
(повышенная активация), высо-кая (в пределах верхней половины зоны
нормы) функциональная активность щитовидной, половых желез и органов
тимико-лимфатической системы. Однако эти изменения наибо-лее
благоприятны при развитии реакции активации на высоких уровнях
реактивности (малая абсолютная величина действующего фактора) и менее
благоприятны (с большей долей глюкокортикоидов и меньшей —
гормонов щитовидной железы и половых желез, а также меньшей стимуляцией
тимико-лимфатической системы) при реакции активации низких уровней
реактивности.
То же самое отмечается и при реакции тренировки. Создается впечатление,
что глюкокортикоиды — это гормоны, четко увеличивающиеся при
нарастании абсолютной величины нагрузки, в то время как гормоны
щитовидной железы и половых желез, напро-тив, по мере увеличения
нагрузки уменьшаются.
На низких уровнях реактивности секретируется больше гормонов (в
пределах соот-ветствующей реакции); в реакции участвует больше
структур, больше энергетические траты, т. е. больше структурная и
энергетическая «цена адаптации» (Меерсон Ф. 3., 1986). На
высоких уровнях реактивности увеличивается чувствительность тканей к
гормонам, в реакции участвует меньше структур, энерготраты меньше и
энергия добывается более экономным путем, т. е. «цена
адаптации»
ниже.
Очень важно, что от уровня реактивности (а не только от реакции)
зависит энерго-обеспечение организма. М. Н. Кондрашова с сотр. (1971
— 1987) обнаружила зависимость от количества действующих факторов
развития различных состояний на уровне митохон-дрий, наряду с
известным, активным (описанным Б. Чансом) — гиперактивное и
гиперак-тивное напряжённое (стрессовое), или просто напряженное. Было
также показано, что при
182
мягких воздействиях, вызывающих развитие активации высоких уровней
реактивности, состояние энергетики митохондрий ближе всего к состоянию
«устойчивого неравнове-сия», по Э. Бауэру (1935).
Возникает вопрос о биологической целесообразности реакций на разных
уровнях реактивности. В общем виде можно сказать, что периодическая
система реакций обеспе-чивает тонкое приспособление к небольшим
изменениям. Что же касается отдельных ре-акций, то кажется непонятной
биологическая целесообразность развития стресса на
не-большие по абсолютной величине воздействия.
Мы думаем, что стресс появился еще на стадии одноклеточных организмов
как ре-акция на сильные, повреждающие воздействия для сохранения любой
ценой жизни клет-ки. Реакции тренировки, спокойной активации и
повышенной активации, по-видимому, также возникли еще на уровне
одноклеточного организма как реакции на определенную интенсивность
действующего фактора — слабую, среднюю и пограничную с сильной.
То-гда же закрепилась закономерность развития разных реакций на разную
интенсивность действующего фактора. В процессе эволюции —
развитии сложного многоклеточного ор-ганизма — эта закономерность
сохранилась, но в другом, усложненном виде — в виде
пе-риодической зависимости этих же, основных, реакций от абсолютной
величины дейст-вующего фактора и от относительной — относительно
каждого уровня реактивности.
В процессе усложнения живых систем, увеличения числа иерархических
уровней чувствительность организма повышается и увеличивается роль
согласованности, синхро-низации и слабых, информационных воздействий.
Поэтому стресс высоких уровней реак-тивности, более мягкий стресс,
развивается уже в ответ не на грубое повреждение, а на нарушение связи
и синхронизации деятельности подсистем организма.
Можно думать, что биологическая роль стресса высоких уровней реактивности и заключается в защите от этих нарушений.
Изучая периодическую зависимость развития адаптационных реакций, мы
столк-нулись с таким фактом: очень часто после развития стресса на
каждом уровне реактивно-сти при увеличении дозы на коэффициент реакции
(К) обнаруживалось отсутствие какого-либо заметного ответа, в первую
очередь, по простому показателю реакции — процент-ному содержанию
лимфоцитов в специально подсчитанной лейкоцитарной формуле. Из-менения
же в тимико-лимфатической и эндокринной системе были более
благоприятны, чем при развитии соответствующей реакции на ту же
величину (силу, дозу) действующего фактора. С явлением отсутствия
реакции столкнулись М. А. Уколова, Г. А. Веревкина (1971) при действии
больших, полулетальных доз противоопухолевых
препаратов.
183
Половина животных погибла при изменениях, характерных для глубокого
стресса, а дру-гая половина осталась жить, и в лейкоцитарной формуле не
отмечалось каких-либо изме-нений по сравнению с исходными анализами, а
в органах тимико-лимфатической системы не было отмечено выраженных
признаков повреждения. Такая ареактивность как способ защиты от
экстремального, почти смертельного воздействия была названа М. А.
Уколовой «реакцией блокады». Однако такое отсутствие
реакции периодически повторялось и при действии небольших по
абсолютной величине, не угрожающих жизни факторов: как от-мечалось нами
ранее, наиболее часто — при увеличении силы (дозы) раздражителя,
на К на фоне стресса каждого уровня реактивности (Гаркави Л. X.,
Квакина Е. Б., 1975—1987). Мы предположили, что между
«триадами» реакций каждого уровня реактивности имеется
«зона ареактивности», расположенная на той же
количественной шкале, что и реакции, и с тем же коэффициентом. Однако
дальнейшие исследования показали, что бывают состоя-ния ареактивности и
не только «после стресса», а и при длительном поддержании в
орга-низме реакции активации (особенно повышенной) на высоких уровнях
реактивности (действие малых по абсолютной величине факторов), и при
длительной переактивации и стрессе низких уровней реактивности
(действии больших по абсолютной величине факто-ров — нагрузок и
патологических процессов). Более редко встречалась
«тренировочная» ареактивность. Попытки специально вызвать
определенное состояние ареактивности с помощью использования К (что
успешно применяется для вызова нужной реакции) оказа-лись
малоэффективными.
Обнаружение существенной разницы между спокойной и повышенной
активацией привело к необходимости выделения отдельной реакции
спокойной активации и повы-шенной активации. Вместо «триад»
реакций и «зоны ареактивности» между триадами оказалась
«тетрада» реакций, а зона ареактивности потеряла свое место
на количествен-ной шкале реакций. Стало ясно, что это не
«зона», а система состояний ареактивности: стрессорная
ареактивность, повышенно-активационная ареактивность,
спокойно-активационная ареактивность и тренировочная ареактивность. По
картине лейкоцитарной формулы каждое состояние ареактивности совпадает
с соответствующей реакцией. По изменениям в тимико-лимфатической и
эндокринной системе — это состояния более бла-гоприятные, чем
соответствующая реакция, вызванная тем же по абсолютной величине
раздражителем. Было показано, что в зависимости от уровня реактивности
(абсолютной величины действующего фактора) состояние ареактивности
может быть более благопри-ятным (на высоких уровнях реактивности
— малая величина раздражителя) или менее благоприятным (на
низких уровнях реактивности).
184
Выяснилась также важная характеристика состояний ареактивности — их стой-кость.
Было обнаружено три стойких состояния ареактивности: стойкое состояние
здоро-вья — повышенно-активационная ареактивность высоких уровней
реактивности и два стойких состояния предпатологии и патологии:
стрессорная ареактивность низких уров-ней реактивности и переактивация
низких уровней реактивности.
Таким образом, стало понятно, что регуляция и поддержание гомеостаза в
орга-низме осуществляется и системой реакций, и
«антисистемой» — системой ареактивности. Это шло
вразрез с нашими представлениями о подвижном, основанном на принципе
ус-тойчивого неравновесия Э. Бауэра (1935) приспособлении живого. Такой
путь приспособ-ления казался нам более свойственным неживой природе.
Повышение резистентности ор-ганизма, связанное с состояниями
ареактивности, как при стойком состоянии здоровья, так и при стойких
состояниях болезни, когда организм как бы переставал реагировать на
воздействия (даже довольно большие), хотя и объясняло высокую
резистентность «здоро-вых до предела» и удивительную
(нередко наблюдаемую) довольно высокую резистент-ность тяжело больных
людей, но казалось также присущим скорее неживому, чем живо-му.
Понять, что это не так, что состояния ареактивности, если можно
сказать, еще бо-лее «живые», чем реакции, помог подход к
анализу и реакций, и состояний ареактивности с позиций уже
установившихся представлений об организме как о сложной нелинейной
колебательной системе. Все живые организмы формировались в процессе
эволюции под влиянием действия различных космических и планетарных
факторов, имеющих опреде-ленные ритмические, частотные характеристики.
Известно, что в организме происходит многократное перекодирование
частоты в интенсивность (силу) и наоборот. При вызове адаптационных
реакций с помощью варьирования интенсивности действующего фактора
обнаружилось четкое отличие индивидуальной чувствительности. Силу
(дозу) приходится всегда подбирать индивидуально. Мы предположили, что
поскольку все живые организмы формировались в условиях действия одних и
тех же частот, разницы в индивидуальной чувствительности на действие
частотного компонента действующего фактора не должно быть, так как
индивидуальная чувствительность связана с коэффициентом
перекодирова-ния силы в частоту. Было также высказано предположение,
что каждой адаптационной реакции присущи свои частотные характеристики
на разных иерархических уровнях (Л. X. Гаркави), но это не так заметно,
благодаря «подстройке» ритмов реакции к более дли-тельным
внешним ритмам.
Проведение исследований с использованием магнитного поля
разной частоты и минимальной
«несущей» величины магнитной
185
индукции во многом подтвердили это предположение. Так, удалось найти
частоты акти-вации, стресса и ареактивности. Тренировочной частоты пока
обнаружить не удалось. При этом почти у всех животных (кроме тех,
которые находились, в стойких состояниях) на однократное действие одной
и той же частоты развивалась одна и та же реакция. Увели-чение частоты
играло ту же роль, что и увеличение интенсивности действующего
факто-ра: происходила периодическая смена тетрад адаптационных реакций,
но К оказался другим, еще меньшим. Известные из литературы по
магнитобиологии данные о наличии «частотных окон»»
— высокоэффективных частот при чрезвычайно низкой интенсивно-сти
магнитного поля — некоторые авторы трактуют в плане
периодического возникнове-ния неспецифических адаптационных реакций
организма (Макеев В. Б., Темурьянц Н. А., 1982; Макеев В. Б„
Темурьянц Н. А., Владимирский Б. М. и др., 1984). Длительное,
систе-матическое, ежедневное повторение воздействия одной частоты в
наших опытах приводило к срыву в стресс. Если такое воздействие
повторяли реже — 1 раз в 3 дня и 1 раз в неделю, то такой срыв
наступал позже. Мы связываем это явление с тем, что разным
ие-рархическим уровням организма, как известно, свойственны свои
частотные характеристики. При действии на мозг с помощью одной частоты
резонансные явления разви-ваются на одном иерархическом уровне, что
способствует десинхронизации работы под-систем организма (Барсукова Л.
П., Гаркави Л. X., Котляревская Е. С, Марьянов-ская Г. Я., 1983;
Гаркави Л. X., Квакина Е. Б., 1984, 1985, 1987). Колебательная природа
биологи-ческих объектов относится к их фундаментальным свойствам
(Ухтомский А. А., Гуляев П. И., 1940; Шноль С. Э., 1967, 1983), тесно
связанным с устойчивой неравновесностью, био-логической ритмичностью,
адаптивностью, механизмами гомеостатического управления. Так как
биоритмы живого организма занимают широкий спектр частот, границы
которого различаются примерно на 10 порядков (Моисеева И. И., Сысуев В.
М., 1981), и чем ниже иерархический уровень, тем больше частота
колебаний, то понятно, как велика роль со-гласованности частотных и
фазовых параметров колебательных процессов в организации живых систем.
Такая согласованность, т. е. фактически интеграция частей в единое
целое, осуществляется благодаря резонансным взаимодействиям, приводящим
к синхронизации колебаний. С этих позиций синхронизацию считают одним
из важнейших системообра-зующих факторов на различных иерархических
уровнях живой природы (Гудвин, 1966, 1979; Блехман И. И., 197.1, 1981;
Романовский Ю. М., Степанова Н. В., Чернавский Д. С, 1971; Винер, 1983;
Путилов А. А., 1987). С синхронизацией, по нашему мнению, связан эффект
взаимоСОдействия при формировании функциональных систем, по П. К.
Анохи-ну.
186
Благодаря взаимному сопряжению множества осцилляторов, контролю над
ними ведущего осциллятора, взаимодействию с факторами среды, в здоровом
организме под-держивается согласованность различных процессов —
составляющих гомеостаза (Ашофф Ю., 1984).
Поэтому в дальнейших опытах мы применяли «аккорд» или
«арпеджио» частот, используя разные частоты либо
одновременно (аккорд), либо последовательно (арпед-жио), начиная с
самых низких, адресованных к мозгу, до довольно высоких, адресован-ных,
например, некоторым эндокринным железам (опыты А. И. Шихляровой). При
таком подходе нам удалось получить обнадеживающие результаты: вызвать и
длительно под-держивать в организме у подавляющего большинства животных
(кроме 5 —15%, находя-щихся в стойких состояниях) физиологическую
реакцию активации (спокойную или по-вышенную) высоких уровней
реактивности. Как показали опыты, синхронизировать дея-тельность
основных регуляторных и защитных подсистем организма у крыс с опухолями
даже под влиянием «арпеджио» частот значительно сложнее.
Однако возможность полной регрессии опухолей (30% случаев)
свидетельствует о перспективности такого подхода. При сочетании таких
воздействий с введением противоопухолевого химиопрепарата
(цик-лофосфана) противоопухолевый эффект резко возрастал (до 80%
регрессии опухолей).
Воздействия на одни лишь местные процессы (с помощью сравнительно
высокой частоты) оказались недостаточно эффективными для вызова
гармоничной общей реакции, возможно из-за малой вероятности
синхронизации высокого порядка, когда частота мест-ного осциллятора
превышает частоту основного — циркадианного — ритма
организма на много порядков.
Установлено, что основой организации живой системы являются
околосуточные (циркадианные) ритмы (Питтендрих К., 1964). Поэтому
большой интерес представляет изучение связи временной организации
биологических объектов с неспецифическими адаптационными реакциями
организма, также имеющими околосуточный ритм.
Известно, что биоритмы организма всегда хотя бы немного отличаются от
ритмов окружающей среды (Питтендрих К.,. 1984; Алякринский Б. С,
Степанова С. И., 1986), что имеет приспособительное значение.
Вследствие того, что каждой адаптационной реакции присущи свои
частотные характеристики, подстройка организма к внешним ритмам при
разных реакциях происходит по-разному, и поэтому циркадианные ритмы
разных реакций отличны. Исследования показали, что степень
синхронизации при разных адаптационных реакциях разных уровней
реактивности также различна, что находит отражение уже
в
187
сигнальном показателе реакции — лейкоцитарной формуле
крови. Оптимальной степе-нью синхронизации колебаний обладает реакция
активации высоких уровней реактивно-сти (действие малых по абсолютной
величине раздражителей). Реакции активации, осо-бенно повышенной,
присущи все черты, свойственные (по А. А. Путилову) синхронизи-рованным
состояниям: высокая устойчивость, оптимизация процессов переноса
вещества, энергии и информации. При снижении уровней реактивности
появляются признаки де-синхронизации даже в реакциях активации и
тренировки (в лейкограмме, например, появ-ляются элементы напряжения).
Стресс низких уровней реактивности — это реакция, при которой на
фоне частичной десинхронизации происходит гиперсинхронизация, что
делает эту реакцию тоже высокоустойчивым состоянием, но относящимся уже
к патологии. Ги-персинхронизация свойственна и переактивации —
второму устойчивому патологическому состоянию.
Все это касается роли колебательных процессов в формировании системы адапта-ционных реакций организма.
Как же рассматривать с этих же позиций состояние ареактивности? Нами
была вы-сказана гипотеза (Л. X. Гаркави), что ареактивность — это
фоновое живое состояние. При развитии любой реакции в ней участвует
только часть структурных элементов каждой подсистемы, органа и ткани. В
мозгу, например, не более 1%. А остальные структурные элементы? Живут
и, по существующим представлениям в этом состоянии «покоя»
— фо-новом живом состоянии, накапливают энергию и пластические
материалы. Как это осу-ществляется? По нашим представлениям, с позиции
роли колебательных процессов, час-тота колебаний в состоянии
ареактивности включает в себя все частотные характеристики реакций: и
тренировки, и спокойной активации, и повышенной активации, и стресса, а
кроме того, колебания других характеристик, возникающие с помощью
параметрического резонанса. Поэтому при ареактивности имеется почти
сплошной спектр колебаний, т. е. происходит энергетическая подпитка от
множества факторов среды. Энергетическое пре-имущество, возможно, и
обусловливает стойкость состояний ареактивности и их большую
благоприятность по сравнению с соответствующей реакцией того же уровня
реактивно-сти. Этот большой спектр частот тем не менее не является
хаосом, просто «белым шу-мом». Это кажущийся хаос, который
по современным представлениям, является фактором самоорганизации. Таким
образом, состояние покоя живых систем, фоновое живое состоя-ние —
ареактивность — еще более активно, чем состояние работы —
реакции, но назна-чение их различно. Ареактивность приобретает черты
тренировочной, спокойно-активационной, повышенно-активационной и
стрессорной, если частоты этих реакций на-чинают
преобладать, но не
достигают некоторого «порога»,
когда ареактивность
188
переходит в реакцию. Такое представление позволяет понять, что
воздействие, вызываю-щее любую адаптационную реакцию (особенно
физиологическую, хорошо синхронизиро-ванную), резонансно подпитывает
энергией фоновое живое состояние. Это совпадает с представлениями М. Н.
Кондрашовой (1979) о положительной роли малых возмущений в процессах
энергетики. Возможно, что и космические воздействия различных частот
резо-нансно подпитывают организм энергией. В связи с этим интересна
роль биологически-активных точек (БАТ) как своеобразных антенн
внутренних органов, таким образом пред-ставленных на поверхности тела.
Во всяком случае, уже установлены факты влияния низ-кочастотных
электромагнитных колебаний естественного происхождения на БАТ
(Нечуш-кин А. И., 1987) и роль при этом их частотно-избирательных
свойств (Нечушкин А. И., Николаев Н. А., 1986). Наши, хотя еще
немногочисленные, исследования говорят об осо-бой роли БАТ в
формировании как реакций на разных уровнях реактивности, так и
со-стояний ареактивности, в том числе стойких. Это согласуется с
имеющимися представле-ниями о рефлексотерапии как о средстве
адаптивного управления (Нечушкин А. И., Нико-лаев Н. А., 1986).
Понимание того, что система адаптационных реакций и система
ареактивности имеют разную шкалу отсчета интенсивности и частоты,
побудило нас искать геометриче-ский образ, соответствующий нашим
представлениям и позволяющий объяснить полу-ченные факты с позиции
представлений об организме как о сложной колебательной сис-теме с
определенными свойствами. Таким оказался образ двойной спирали
(предложен Л. X. Гаркави), состоящей из двух коаксиально расположенных
расширяющихся спиралей. Системе адаптационных реакций соответствует
внутренняя (более узкая) спираль, а сис-теме ареактивности —
внешняя (более широкая). Этот образ отражает периодичность, цикличность
смены реакций и состояний ареактивности (развитие их на разных витках
спирали), наличие определенного коэффициента и наличие экспоненциальной
зависимо-сти между интенсивностью (частотой) действующего фактора и
характером ответной ре-акции.
Анализ трех устойчивых состояний (при реакции повышенной активации
высоких уровней реактивности, стрессе и переактивации — низких
уровней реактивности) показы-вает, что во всех этих случаях
устойчивость — неизменяемость гомеостатических пара-метров
— не только при малых, но и при сравнительно больших воздействиях
связана с резонансным увеличением амплитуды колебаний, что приводит к
переходу со «спирали реакций» на «спираль
ареактивности» (Гаркави Л. X., Квакина Е. Б., Серегин Ю. В.,
Фай-дыш Е. А., 1985).
Если образ расширяющейся «двойной спирали» справедлив и для адаптационной
189
деятельности подсистем организма, органов и тканей,
то при хорошей синхронизации работы подсистем организма на разных
иерархических уровнях образ колебательной рит-мической деятельности
всего организма представляет собой симметричную, стройную картину.
Однако реальный «образ» такой деятельности организма всегда
отличен и инди-видуален, что соответствует представлениям А. П. Дуброва
(1987) о существовании мно-гообразных LDS-реакций. Вообще тот факт, что
на одну и ту же силу раздражителя раз-личные организмы отвечают разными
адаптационными реакциями, отражает наличие, по А. П. Дуброву,
функциональной биосимметрии в живых организмах. Существование
мно-гомерных силовых (частотных) отношений в организме и системы
стандартных реакций, зависящих от абсолютной и относительной величины
действующего фактора и исходного состояния, никоим образом не
противоречит представлениям об индивидуальных различиях реакций,
вероятность которых тем больше, чем более многомерны отношения как
внутри организма, так и его взаимоотношения с внешними факторами.
С различными частотными характеристиками реакций и ареактивности мы
связы-ваем и разную скорость течения биологического времени, и
возможность его обратного направления — против стрелы времени. И.
А. Аршавский (1979) связывает такую воз-можность с фазой избыточного
анаболизма, что, в свою очередь, приводит к увеличению негэнтропии. По
сути, мы стоим на тех же позициях. Энергетические траты и процессы
анаболизма при реакциях на высоких уровнях реактивности (малая сила и
частота дейст-вующих факторов) более благоприятны, а особенно
«анаболическая» реакция— реакция с избыточным
анаболизмом - это реакция повышенной активации высоких уровней
реак-тивности. Мы думаем, что эта реакция на каких-то уровнях
реактивности: соответствует «физиологическому стрессу»,
найденному и описанному Аршавским.
Известно, что при старении и болезнях биологическое время течет быстрее
(Мои-сеева И. И., Сысуев В. М., 1981; Аршавский И. А., 1982), чем в
молодости и у здоровых. Нами показано, что по мере старения (и при
развитии заболеваний) организм стремится на низкие уровни реактивности
с большей частотой колебательных процессов. И напротив, у молодых и
здоровых реакции развиваются на высоких уровнях реактивности с меньшей
частотой колебательных процессов. С представлениями о скорости и
направленности био-логического времени совпадают и представления об
энтропии и негэнтропии живого ор-ганизма. Признавая важную роль
специфических механизмов в учении о детерминанте Г. Н. Крыжановского
(1980), мы думаем, что в переходе нормальной детерминанты в
пато-логическую играет существенную роль увеличение частотных
характеристик, благодаря сильному воздействию какого-либо агента на
область, вызывающую формирование де-терминанты.
190
Изучение системы адаптационных реакций и состояний ареактивности по
мере старения (и при развитии главных неинфекционных болезней)
показало, что при этом происходит переход на высокие этажи (низкие
уровни реактивности), реакции и состоя-ния ареактивности на которых
характеризуются, как уже говорилось, все большим откло-нением
гомеостаза от уровня нормы молодого здорового организма. Это полностью
со-гласуется с «законом отклонения гомеостаза», открытым В.
М. Дильманом (1978, 1987).
Необходимо отметить, что если в организме спонтанно (или
целенаправленно) при старении поддерживается реакция активации, лучше
на высоких уровнях реактивности (но даже и на довольно низких),
отклонение гомеостаза происходит значительно медлен-нее и процессы
старения также замедляются. На фоне же преобладания стресса, особенно
тяжелого стресса низких уровней реактивности, наблюдается отклонение
гомеостаза, ве-дущее к быстрому старению. Мы считаем, что система
неспецифических адаптационных реакций имеет прямое отношение и к
механизмам старения, и к механизмам антистарения.
Система ареактивности также имеет отношение к этим механизмам. В
молодости (у здоровых) чаще всего имеется состояние стойкой
повышенно-активационной ареактив-ности, при котором наиболее выражен
принцип неравновесности Э. Бауэра. Оно устойчи-во, благодаря хорошей
синхронизации, увеличению вследствие этого амплитуды колеба-ний,
приводящим к частым «перескокам» со «спирали
реакций» на «спираль ареактивно-сти» и наоборот. В
старости же, особенно глубокой, когда вместо повышения секреции
центральных и периферических гормонов в начальные периоды старения
(Дильман В. М., 1978, 1987), отмечается, напротив, их снижение,
особенно часто встречается ареактив-ность низких уровней реактивности.
Она более благоприятна, чем стойкие патологические состояния, хотя и
существенно отличается от нормы молодого здорового организма. Од-нако
именно с развитием такого состояния ареактивности, более
благоприятного, чем ре-акция соответствующего уровня реактивности, мы
связываем довольно высокую рези-стентность организма, которая нередко
наблюдается у людей большого возраста.
В заключение мы хотим перечислить основные принципы регуляции
гомеостаза на основе периодической количественно-качественной
закономерности развития системы адаптационных реакций и ареактивности.
1. В регуляции гомеостаза существенную роль играет
как система различных не-специфических адаптационных реакций организма
— реакции тренировки,
спокойной и повышенной активации,
стресса, так и соответствующие реакциям состояния ареак-тивности.
2. Развитие адаптационных реакций подчиняется периодической
(циклической) за-кономерности: по мере увеличения абсолютной величины
(силы, дозы) действующего
191
фактора происходит многократное повторение основных реакций на разных
уровнях реак-тивности организма. С этим связано наличие многих
оптимумов в живых системах. Коли-чественные переходы дискретны и имеют
определенный коэффициент. Эта зависимость является логарифмической.
3. Силовые отношения в организме многомерны. Абсолютная величина
действую-щего фактора определяет уровень реактивности, на котором
развиваются реакции: на ма-лую абсолютную величину— высокие
уровни, на большую — низкие. Относительная ве-личина действующего
фактора (относительно данного уровня) определяет тип реакции на каждом
уровне реактивности: слабая относительная величина — реакцию
тренировки, средняя?— спокойную и повышенную активацию, большая
— стресс. Состояния ареак-тивности имеют иную количественную
шкалу.
4. По мере старения или при тяжелых заболеваниях организм мужчин
постепенно теряет способность реагировать на малые по абсолютной
величине действующие факто-ры. Женщины любого возраста, за редким
исключением, эту способность сохраняют.
При сохранении способности реагировать на малые возмущения факторы,
малые по абсо-лютной величине, могут обладать более эффективным
действием, т. е. в живых системах работает принцип минимизации.
Способность реагировать на малые воздействия можно вернуть, используя
способы вызова реакции активации (особенно повышенной) высоких уровней
реактивности.
5. При длительном поддержании реакций активации и тренировки высоких
уровней реактивности, особенно повышенной активации (у молодых,
здоровых без специальных воздействий или при длительных
целенаправленных активирующих воздействиях) орга-низм способен
«выбирать» из числа падающих раздражителей малые по
абсолютной ве-личине. При длительном поддержании стресса, переактивации
и напряженных реакций активации и тренировки на низких уровнях
реактивности (вследствие болезни, перегру-зок, старения) организм
«выбирает» большие по абсолютной величине раздражители.
6. Эффективность раздражителя для вызова реакции тем выше,
чем большей но-визной «по качеству» или «по
количеству» он обладает.
7. Мощность развивающихся адаптационных реакций на разных уровнях
реактив-ности может поддерживаться и с помощью разного участия
функциональных структур (большего на низких уровнях), и
с помощью разной чувствительности к
биологически-активным веществам, и с помощью качественно отличного
функционирования структур в зависимости от типа реакций, а также разным
энергетическим обеспечением.
8. Состояния ареактивности
характеризуются отсутствием видимой
реакции, стойкостью, более
благоприятными изменениями
192
в организме, чем при соответствующей адаптационной реакции на ту же
абсолютную ве-личину раздражителя, и существенной ролью в их развитии
фактора повторности дейст-вия раздражителя. Они также подчиняются
периодической количественно-качественной закономерности.
9. Каждую адаптационную реакцию характеризует свой спектр частот
на разных иерархических уровнях организма. Частоты реакций
возрастают по мере
увеличения абсолютной величины
действующего фактора. С этим связана роль резонанса и синхро-низации в
механизме развития разных адаптационных реакций.
10. Соответствующие реакциям состояния ареактивности,
являющиеся фоновым живым состоянием
— аналогом состояния покоя, характеризуются
спектром частот, включающим как частотные характеристики реакций, так и
колебания других параметров, возникающие с помощью параметрического
резонанса. Такое представление объясняет стойкость состояний
ареактивности и возможность осуществления
резонансного обога-щения живых
систем энергией. С этой точки зрения, состояние
покоя (ареактивности) живых систем даже более активно, чем
состояние работы (реакции), но назначение его иное.
11. Неспецифическая резистентность,
связанная с неспецифическими адапта-ционными
реакциями, может быть активной и пассивной. Активная
резистентность реализуется благодаря высокой
функциональной активности защитных систем организма и преобладанию в
мозгу физиологических процессов возбуждения. Такая
резистентность характеризует реакцию, активации, особенно
повышенную, высоких уровней реактивно-сти. Пассивная резистентность
реализуется благодаря преобладанию
в мозгу процес-сов
охранительного (реакция тренировки) или запредельного
(стресс) торможения,
обеспечивающего снижение
возбудимости, чувствительности.
12. С состоянием ареактивности связаны два других типа
резистентности: «под-вижная» и «жесткая».
«Подвижный» тип резистентности обусловлен частыми
резонанс-ными переходами между состояниями
повышенно-активационной ареактивности и
стойкой реакцией повышенной активации высоких уровней
реактивности. Подвижная резистентность поддерживает гомеостаз на
физиологическом уровне. «Жесткая»
рези-стентность обусловлена только состоянием ареактивности в
результате длительного тяже-лого стресса, а также переактивации низких
уровней реактивности и поддерживает гомеостаз за пределами
физиологических значений.
13. Степень неравновесности и величина энтропии живой
системы зависят как от
характера реакции или
состояния ареактивности, так и от уровня, на котором они
193
развиваются. Наиболее неравновесными состояниями с высокой негэнтропией
являются стойкая повышенная активация высоких уровней реактивности и
соответствующая ей ареактивность. Наиболее близким к
термодинамически равновесному
состоянию с максимальным
производством энтропии является стресс низких уровней
реактивности.
ЗАКЛЮЧЕНИЕ
Описанные в книге система
неспецифических адаптационных реакций — об-щих и
местных — и система ареактивности, развитие
которых подчиняется периоди-ческой
количественно-качественной закономерности, являются реальной основой
разви-тия в организме различных дискретных состояний, реальным
механизмом многоуровневой регуляции гомеостаза. В зависимости от
абсолютной и относительной величины действующего, управляющего фактора
и исходного состояния развиваются качественно отличные адаптационные
реакции или состояния ареактивности. В каждый данный мо-мент в
организме имеются какая-то реакция или состояние, характеризующиеся
комплек-сом изменений, определяемым типом адаптационной реакции или
состоянием ареактив-ности и уровнем реактивности, на котором они
развиваются.
Если реакции спокойной и повышенной активации или тренировки
развиваются на высоких уровнях реактивности (когда организм способен
выбирать для реагирования ма-лые по абсолютной величине действующие
факторы) или имеется состояние повышенно-активационной ареактивности,
то это означает, что уровень гомеостаза соответствует норме, здоровью.
Как видно, состояние здоровья не одно — существует довольно много
различных «состояний здоровья», что соответствует принципу
многооптимумости и сов-падает с представлениями еще великих ученых
древности.
Наиболее стойким состоянием здоровья с характером изменений в
организме, со-ответствующим верхним границам нормы, близкой к норме
молодого, здорового организ-ма, является состояние
повышенно-активационной ареактивности. Тот же характер изме-нений, но
менее стойко поддерживаемый, отмечается при развитии реакции повышенной
активации. При развитии реакции спокойной активации характер изменений
близок к верхней половине зоны нормы, но функциональная активность
защитных и регуляторных систем несколько ниже и соотношение активности
отдельных подсистем организма не-сколько иное. При развитии реакции
тренировки также высоких уровней реактивности характер изменений близок
к нижней половине зоны нормы и устойчивость этого состоя-ния меньше.
Если реакции тренировки, спокойной и повышенной активации (а также
состояния ареактивности) развиваются на низких уровнях реактивности
(когда организм способен выбирать для реагирования довольно большие
действующие факторы или действие этих
194
факторов продолжается долго), это соответствует также множеству
состояний, промежу-точных между здоровьем и болезнью. В эту же группу
состояний входит мягкий стресс — стресс высоких уровней
реактивности. Тяжесть «промежуточных» состояний и их
харак-тер связаны с уровнем реактивности и наличием в организме
«слабого звена» — какой-либо наиболее уязвимой
подсистемы, которая определяет специфику предболезни.
Если реакции стресс и переактивация развиваются на очень низких уровнях
реактивности, а реакции тренировки и активации становятся резко
напряженными и рассогласованными (что также говорит о вынужденном
реагировании на очень большие действующие факто-ры), то эти состояния
являются неспецифической основой различных патологических процессов.
При многократном или систематическом действии сильных раздражителей
развиваются особенно стойкие патологические состояния: стрессорной и
переактивацион-ной ареактивности. Характер заболеваний, по нашему
мнению, определяется в значитель-ной мере типом преобладающей в данном
организме, у данного индивидуума, адаптаци-онной реакции, что,
возможно, закреплено генетически. Иными словами, это связано с
конституцией человека, во многом определяющей в разных условиях жизни
тактику и стратегию адаптации, поддержания необходимого для жизни и, по
возможности, опти-мального для данных условий уровня гомеостаза.
Выделение в качестве третьего главного признака конституции (кроме
продольных размеров тела и доминирующего типа обмена веществ) типа
реагирования (Казначеев В. П., Казначеев С. В., 1986) совпадает с
нашими представлениями о значении характера преобладающей адаптационной
реакции для формирования конституционных особенностей.
Эти представления ни в коем случае не отрицают наличия специфики
действую-щих факторов и различных болезней. Нужно только помнить, что
каждый действующий фактор обладает и качеством, определяющим специфику
его действия, и количеством, оп-ределяющим неспецифическую сторону его
действия, и, следовательно, характер разви-вающейся неспецифической
реакции. Специфические изменения накладываются на об-щий
неспецифический фон. Наши исследования, посвященные, главным образом,
изуче-нию роли неспецифических адаптационных механизмов, говорят об их
значении для фор-мирования основы состояний здоровья, предболезни и
болезни, а также старения.
Возможность подойти к управлению этими состояниями на основании
обнаруже-ния периодической количественно-качественной закономерности их
развития открывает перспективы использования целенаправленного вызова
«состояний здоровья» в практике для оздоровления,
профилактики и повышения эффективности лечения.
Уже на данном уровне развития исследований можно
предложить для этого теоретиче-ски обоснованную
систему мероприятий на разных этапах активной профилактики
и
195
лечения, с учетом пола, возраста и индивидуальной чувствительности.
Советская медици-на с самого начала своего существования была
направлена на активную профилактику. Это помогло преодолеть большинство
массовых инфекций, которые в то время являлись основной причиной смерти
от болезней. В настоящее время на первый план выступили неинфекционные
болезни, по отношению к которым в медицине еще не разработана
эф-фективная система активной профилактики. Основные мероприятия и
основные средства направлены на лечение и затем — диспансеризацию
обширного контингента хронических больных. Широкое (и часто
недостаточно обоснованное) применение антибиотиков, глю-кокортикоидных
гормонов и различных синтетических лекарственных препаратов,
подав-ляющих активность защитных систем организма, приводит к быстрому
«подлечиванию» заболеваний, выведению из обострения
болезни, но способствует увеличению числа хро-нических больных. Особое
значение при этом имеет применение больших доз (силы) ле-чебных средств
без достаточно обоснованного индивидуального подбора величины
дейст-вующего фактора.
С философских позиций это объясняют тем, что в центре представлений
медицины находится категория «болезнь», а не категория
«здоровье» (Щепин О. П., Царегородцев И. Г., Ерохин В. Г.,
1983; Ерохин В. Г., 1985). Поэтому в настоящее время необходима
перестройка советской медицины в плане настоящей активной профилактики,
которая ставит во главу угла категорию «здоровье» и которую
необходимо проводить не тогда, когда бо-лезнь уже развилась и процессы
старения зашли далеко, а при малейших отклонениях от оптимального
уровня здоровья.
Когда же нужно начинать проведение профилактических мероприятий? В. М.
Дильман (1968—19870) считает, что это нужно делать уже в
20—25 лет, когда найденный им закон отклонения гомеостаза
(связанный с ростом и развитием организма) начинает работать, отклоняя
гомеостаз организма все дальше от нормальных значений и способст-вуя
развитию главных неинфекционных болезней и старения. И. А. Аршавский
(1958—1985), изучая организм в процессе онтогенеза, говорит о
большом числе физиологически незрелых младенцев (что в дальнейшем
приводит к высокой заболеваемости) и предлагает комплекс нормализующих,
повышающих резистентность организма воздействий, на осно-ве своих
теоретических исследований о роли умеренной двигательной активности,
гипок-сии и холодового закаливания. Исходя из его представлений,
активную профилактику нужно начинать буквально с младенчества (при
наличии показаний).
Мы на основании проведенных исследований считаем, что в наше время, когда
196
распространены гипокинезия и гиподинамия, наряду с избыточным и часто
нерациональ-ным питанием, избыточным употреблением спиртных напитков,
фармакологических пре-паратов, большими психоэмоциональными нагрузками
и обширной (не всегда полезной) информацией, при наличии
неблагоприятной экологической обстановки, профилактиче-ские мероприятия
нужно начинать как можно раньше, также с учетом показаний.
Обсле-дование детей, подростков, девушек и юношей показало, что у
многих есть отклонения от «нормы» адаптационных реакций,
даже при отсутствии определенных заболеваний. Если не вернуть их в
«границы нормы», то можно считать (с большой долей
вероятности), что в дальнейшем у них разовьется какая-либо предболезнь,
а затем и болезнь.
Очень важны критерии риска — перехода от нормы в промежуточные
состояния между здоровьем и болезнями. Такие критерии предлагают многие
ученые: для раннего детства, как уже говорилось, — И. А.
Аршавский, после 20—25 лет — В. М. Дильман; глубокие
исследования промежуточных состояний между здоровьем и болезнью
проводит Р. М. Баевский (1979), который предлагает интересный
показатель оценки состояния по анализу частоты сердечных сокращений
— ЧСС, свидетельствующий о разной степени нарушения нормы. Наши
предварительные исследования и исследования С. 3. Клецкина (1983)
говорят о хорошем совпадении изменений «показателя ЧСС» с
показателями раз-личных адаптационных реакций. Мы предлагаем простой
показатель реакций — тщательный (на 200 — 300 клеток)
подсчет лейкоцитарной формулы, в которой процент лимфо-цитов определяет
тип адаптационной реакции, а отсутствие или наличие отклонений от нормы
других элементов лейкоцитарной формулы (моноцитов, эозинофилов,
палочкоя-дерных нейтрофилов и базофилов), а также лейкоцитоза или
лейкопении — соответствен-но отсутствие или наличие признаков
напряженности реакций. Признаки напряженности — причем имеет
значение их число и степень их выраженности — свидетельствуют о
низких уровнях реактивности (высокие этажи), на которых развиваются
адаптационные реакции, т. е. о той или иной степени отклонения от нормы
даже при наличии реакций тренировки или активации.
Не нужно забывать, уповая только на сложные показатели лабораторных и
аппара-турных исследований, такие простые, показатели, как
самочувствие, работоспособность, настроение, заболеваемость в течение
какого-то отрезка времени. При наличии какого-либо, даже минимального
отклонения от здоровья, нормы, мы предлагаем широко прово-дить лечение,
которое мы назвали «диагностическим». Давно известно
применение для дифференциальной диагностики лечения ex juvantibus. Всех
людей, кто предъявляет ка-кие-то жалобы или имеет отклонение от нормы
по каким-то критериям, нужно лечить. Поэтому мы предлагаем для
диагностического лечения простой вариант активационной
197
терапии малыми дозировками природных средств с сочетанием общего и
местного воз-действия. Этот способ безвреден, оказывает быстрый эффект
только в случаях наличия промежуточных состояний между здоровьем и
болезнью или самых начальных стадиях заболевания и тем самым позволяет
выделять группы риска.
Как уже неоднократно говорилось, для активационных профилактических
меро-приятий мы широко применяем биостимуляторы растительного и
животного происхожде-ния, чаще всего элеутерококк (Брехман И. И.,
1968). Одновременно с приемом «управ-ляющих» активационных
средств при активационной терапии применяется биохимиче-ское
сопровождение (термин М. Н. Кондрашовой) в виде добавления недостающих
суб-стратов (метаболитов, гормонов, витаминов, микроэлементов и т. д.,
и т. п.). Прекрасным средством как активационного воздействия, так и
биохимического сопровождения при самых различных заболеваниях (в первую
очередь — ишемической болезни сердца) явля-ется, по нашим данным,
янтарная кислота (и ее соли, особенно соли калия), важная роль которой
в энергетическом обмене показана многолетними исследованиями
Кондрашовой (1968 — 1985).
Необходимо еще раз подчеркнуть, что мы считаем важным залогом успеха
различ-ных профилактических и лечебных мероприятий четкое и хорошо
организованное выпол-нение принципа этапности.
С целью повышения уровня здоровья можно использовать различные способы
вы-зова и поддержания в организме физиологической адаптационной реакции
активации, по-вышающей сопротивляемость организма, и борьбы со
стрессом, переактивацией и напря-женными реакциями тренировки и
активации на разных этапах профилактики, диспансе-ризации и лечения.
Эти способы просты, доступны для каждого, не требуют анализов крови и
высоко эффективны.
1. Для активной профилактики заболеваний и повышения уровня здоровья у
«прак-тически здоровых» людей необходимо проводить
периодические курсы активационной профилактической терапии с
использованием специального режима, основанного на принципах новизны и
минимизации воздействий (режим № 1) с учетом пола и возраста. Эти курсы
нужно проводить особенно весной и осенью, в период наиболее высокой
забо-леваемости. Исследования показали, что применение таких режимов
существенно (на 30 — 50%) снижает заболеваемость, значительно
улучшает работоспособность, настроение и самочувствие. В случае же
заболевания оно протекает легче и выздоровление наступает быстрее. Этот
режим прост, не требует проведения анализов крови и высоко эффективен
для людей любого пола и возраста, особенно для женщин, детей и
стариков. У здоровых при исследовании лейкоцитарной формулы обычно
отмечаются либо реакции активации (спокойная или
повышенная) без
198
элементов напряженности, либо с наличием нерезко выраженных и нестойких
элементов напряженности. После использования режима преобладает реакция
активации без выра-женных элементов напряженности.
2. Разработан режим и для активной профилактики заболеваний
у людей в «треть-ем» состоянии (предболезни,
донозологическом состоянии), в котором
находится от 50 до 80%
населения (Баевский Р. М., 1979). Эти люди предъявляют
жалобы общего характера, иногда
преобладают жалобы,
свидетельствующие о наличии
«местной» патологии (боли в области желудка, кишечника,
сердца и т. п.). При преобладании
жа-лоб общего характера нужно вызвать реакцию
активации также с помощью режима оз-доровления, основанного на
использовании принципов новизны и минимизации (режим № 1).
При более четкой выраженности «местных» жалоб можно
применять режим № 2, основанный на знании биоритмических особенностей
адаптационных реакций (нервной и гуморальной фазы), а также разной
чувствительности иерархических уровней организма. Он может быть
использован при необходимости сочетания местного и общего действия. Он
также прост, доступен, высоко эффективен.
3. Для лечения и оздоровления при хронических заболеваниях,
длительном дейст-вии больших нагрузок или повреждающих факторов (в том
числе, так называемых лечеб-ных) разработан режим № 3, основанный на
экспоненциальной зависимости между дозой (силой) действующего фактора и
характером реакции, а также принципе минимизации. Хорошие
результаты наблюдались более чем в 70% случаев. Этот режим также прост,
не требует анализов крови и высоко эффективен, особенно у людей
пожилого возраста и да-же старых, хорошо сочетается с любым специальным
лечением, повышая его эффектив-ность и снижая побочное действие.
4. Для лечения начальных (функциональных) стадий
заболевания и одновре-менно
дифференциальной диагностики, выявления
«факторов риска» проводится так называемое
«диагностическое лечение».
Поскольку диагностика не является самоцелью, а лишь средством для
улучшения лечения, а в начальных, функциональных стадиях заболевания
обычные диагностические методики не информативны и не безразличны для
организма, то целесообразно объеди-нить диагностику и лечение.
Проведение диагностического лечения стало реальным бла-годаря
существенной разнице в эффективности активационной терапии: быстро
насту-пающего эффекта при начальных стадиях заболеваний и медленно
— при тяжелых забо-леваниях со значительными структурными
нарушениями.
Вместо дорогостоящих и не всегда безвредных диагностических мероприятий
больным в течение 3—4-х недель
проводят активационную терапию с использованием
199
режима № 1 или режима № 2 с обязательным сочетанием общего и местного
действия. Если по истечении 3—4-х недель такого лечения
происходит существенное улучшение (уменьшение или исчезновение жалоб),
это свидетельствует о том, что заболевание функ-циональное, в начальной
стадии. Проведение диагностических мероприятий в этой стадии
малоэффективно, а лечить таких больных необходимо, чтобы болезнь не
стала тяжелой и хронической.
При отсутствии существенного улучшения больной должен быть обследован
для установления диагноза и дальнейшего лечения. Исследования показали,
что таких боль-ных обычно остается не более 20%.
5. В стационаре для
повышения неспецифической резистентности на
фоне интенсивной специфической терапии желательно осуществлять подбор
доз действующих активирующих средств по специально подсчитанной
лейкоцитарной формуле — простому и информативному показателю
реакций, т. е. с использованием кибернетического прин-ципа
управления с обратной связью.
Это относится и к
онкологическим больным. При
невозможности производства анализов можно использовать
режим № 2 или режим № 3. После выздоровления или между курсами лечения
можно переходить на режим № 1.
6. Для повышения эффективности активационной профилактики и
терапии у всех людей (кроме больных,
которым это в данное время противопоказано) необходимо со-четать
ее с каким-либо циклическим видом мышечного движения
(ходьба, бег, плавание, лыжи, гимнастика). Если дозы активирующих
воздействий в процессе проведения профи-лактики и лечения нужно
стремиться снижать, то ритмическое мышечное движение
не-обходимо постепенно увеличивать (до индивидуального оптимума).
7. На санаторно-курортном этапе лечения и оздоровления
также необходимо про-ведение активационного лечения по одному из
режимов. Санаторно-курортные факторы по своей природе являются
активационными, но обычно избыточными по количеству, что часто приводит
к обострениям заболеваний и переходу в стресс.
Активационная терапия способна
умерить слишком сильное
действие этих факторов и тем повысить эффек-тивность
лечения.
Исследования, проведенные в различных санаториях, показали, что лишь у
не-большой части больных стойко держится реакция активации или
тренировки. У осталь-ных происходит чередование различных реакций, в
том числе, часто встречается стресс. У некоторых стресс является
преобладающей реакцией. Таким образом, даже при правиль-ном проведении
специального лечения основная задача санаторно-курортного лечения
— оздоровить человека, повысить его
устойчивость, сопротивляемость, выполняется
да-леко не
200
всегда. Действительно, если часть людей после санаторно-курортного
лечения получает заряд на весь год, то у многих существенного,
длительного улучшения не отмечается, а у некоторых даже наблюдается
ухудшение. Мы связываем это именно с тем, что санаторно-курортное
лечение не использует целенаправленного повышения резистентности с
помо-щью развития адаптационной реакции активации.
8. Активационную терапию (по режиму № 1) необходимо проводить
беременным до достижения донашиваемости плода,
лечения токсикозов и получения физиологически зрелых, полноценных
плодов. Результаты показали высокую эффективность (более 80%) режима в
этих случаях.
9. Активационную терапию по одному из режимов необходимо
проводить детям, подросткам, юношам и девушкам для их нормального
развития и повышения
сопро-тивляемости организма. Дети, подростки,
юноши и девушки легко поддаются актива-ционной терапии, так как реакция
активации свойственна людям этого возраста. Оздо-ровление в
юном возрасте является условием здоровья и
хорошей работоспособности в зрелые годы.
10. Активационную терапию по одному из режимов необходимо
проводить лицам пожилого возраста для замедления темпов старения и
борьбы с преждевременным старе-нием.
Наблюдения показали, что активационные курсы лечения лучше проводить не
один, а два, иногда даже три раза в год, например, в условиях
профилакториев или амбу-латорно. Такие активационные курсы могут быть
проведены без отрыва от производства.
Использование открытых в нашей стране физиологических адаптационных
реакций для повышения уровня здоровья и активной профилактики
(диспансеризации) будет особенно эффективно при создании строгой
этапности мер профилактики и лечения. Предлагаемая система
оздоровительных мероприятий на разных этапах активной профилактики и
лече-ния не требует больших затрат ввиду использования малых
(значительно меньших тера-певтических) доз природных биостимуляторов
без применения каких-либо дорогостоящих методов диагностики и контроля
и поэтому пригодна для массового применения.
Для реального осуществления предлагаемых мероприятий, научно
обоснованных многолетними исследованиями по разработке и развитию
открытия № 158 (как нами, так и нашими последователями во многих
городах Советского Союза), необходимо создание учебного центра или
постоянно действующего семинара для врачей, в первую очередь общего
профиля (участковых, в будущем семейных врачей), а также для врачей
разных специальностей.
Хорошая организация теоретически обоснованной активационной терапии
должна способствовать оздоровлению
больших контингентов населения и является вполне
201
реальной в настоящее время. Хочется еще раз подчеркнуть, что хотя на
человека в течение жизни падают различные нагрузки, действуют самые
различные повреждающие факторы, но от уровня неспецифической
резистентности (устойчивости, сопротивляемости) целого организма во
многом зависит и переносимость нагрузок, и защита от повреждающего
дей-ствия, и заболеваемость, и переносимость болезни, и скорость, и
полнота выздоровления. Поэтому мы убеждены, что дальнейшее развитие и
совершенствование управления неспе-цифической резистентностью организма
с помощью целенаправленного развития в нем необходимой общей
адаптационной реакции приведет к расширению границ использова-ния этих
реакций в целях массового оздоровления населения.
202
ЛИТЕРАТУРА
Акоев И. Г. Некоторые итоги и очередные задачи электромагнитобиологии//
Проблемы экспери-ментальной и практической электромагнитобиологии.
Пущино, 1983. С. 1-27.
Акоев И. Г. Принципиальные особенности изучения биологической опасности
и нормирования электромагнитных излучений//Биологические эффекты
электромагнитных полей. Вопросы их ис-пользования и нормирования.
Пущино, 1986. С. 124 — 129.
Аладжалова Н. А. Медленные электрические процессы в головном мозге. М., 1962. 240 с.
Александровская М. М. Приспособительные механизмы в центральной нервной
системе//Архив патологии. 1968. Т. 30. № 8. С. 26—32.
Алешин Б. В. Гистофизиология гипоталамо-гипофизарной системы. М., 1971." 437 с.
Алешин Б. В. Эндокринная система и гомеостаз//Гомеостаз. М., 1976. С. 60—85.
Алякринский Б. С, Степанова С. И. По закону ритма. М., 1985. 176 с.
Анохин П. К. Очерки физиологии
функциональных систем. М.,
1975. 306 с.
Апшкалне Д. Л., Лиепа М. Э., Кикут Р. П. Эффект постоянного магнитного
поля (ПМП) на синтез ДНК глиом человека in vitro.//Биологическое
действие электромагнитных полей. Тезисы докл. Всесоюз. симпозиума.
Пущино, 1982. С. 77 — 78.
Апшкалне Д. Л., Лиепа М. Э., Кикут Р. П. и др. Индивидуальная
чувствительность глиом человека к разным биологически активным
агентам//Магнитные поля в биологии, медицине и сельском хо-зяйстве.
Тезисы докл. Ростов н/Д, 1985. С. 69-71.
Арефьева Л. М. Холинэстереза мозга и крови при изменении резистентности
организма с помо-щью воздействия на гипоталамус: Автореф. дис. ...
канд. биол. наук. Ростов н/Д, 1972. 21 с.
Аршавский И. А. Особенности адаптации и механизмы ее осуществления в
различные возрастные периоды//Приспособительные возможности стареющего
организма. Киев, 1968. С. 61—78.
Аршавский И. А. Биологические и медицинские аспекты проблемы адаптации
и стресс в свете данных физиологии онтогенеза//Актуальные вопросы
современной физиологии. М., 1976. С. 144—191.
Аршавский И. А. Физиологические механизмы и закономерности индивидуального развития. М., 1982. С. 270.
Аршавский И. А. Некоторые методологические и теоретические аспекты
анализа закономерностей индивидуального развития организмов//Вопросы
философии. 1986. № 11. С. 95-104.
А. с. 1194419 (СССР), МКИ4 А 61 К 31/00. Способ лечения хронических не
специфических саль-нингоофоритов/0. Л. Деев, JI. М. Мелехина, Д. Ю.
Мирович. (СССР). № 3617547/28-14; Заявл. 18.05.83; Опубл.
30.11.85//Бюл. Открытия. Изобретения. 1983. № 44.
А. с. 995792 (СССР), МКИ3 А 61 К 35/26, А 61 К 37/24. Способ лечения
психических заболева-ний/А. И. Шевелев, М. П. Невский, Л. X. Гаркави,
М. А. Уколова, Е. Б. Квакина (СССР). № 3317796/28-13; Заявл. 06.07.81;
Опубл. 15.02.83// Бюл. Открытия. Изобретения. 1981. № 6.
А. с. 754726 «Т» (СССР), МКИ2 А 61 В 5/02. Устройство для
расчета доз воздействия при неспе-цифической терапии/Копп И. Г., Пинчук
Н. И., Гаркави Л. X., Квакина Е. Б. (СССР). № 2628320/28-13;
Заявл. 12.06.78.
203
А. с. 522688 (СССР), МКИ2 А 61 № 1/42. Способ лечения злокачественных
опухолей/Панков А. К., Уколова М. А., Квакина Е. В., Гаркави Л. X.,
Бражни-кова Е. И., Салатов Р. Н., Соловьева Г. Р., Еремин В. А. (СССР).
№ 2084728/28-13 А 63 № 1/42; Заявл. 12.12.74. Гриф секретности снят
(письмом Госкомизобре-тений СССР № 284728/13 от 9.02.77).
А. с. 1266520 (СССР), МКИ А 61 В 5/00. Способ определения адекватности
нагрузок при зимнем плавании/Бутов М. А., Ногаллер А. М. (СССР). №
3004243/ 28—14; Заявл. 04.09.80; Онубл. 30Л0.86//Бюл. Открытия.
Изобретения. 1986. № 40.
А. с. 1474925 «Т» (СССР), МКИ4 А С1 К 35/78. Способ
восстановления физической и умственной работоспособности/Гаркави Л. X.,
Квакина Е. Б., Лупи-чев Л. Н., Файдыш Е. А., Кузьменко Т. С. (СССР). №
3759123/28-14; Заявл. 22.05.84.
Африканцева Л. П., Бутов М. А., Денисова А. И. Общие неспецифические
адаптационные реакции организма у больных инфарктом миокарда в процессе
ргабилитации//МРЖ. 1985. № 10227. С. 12.
Ашофф Ю. Свободнотекущие и захваченные циркадианные ритмы//Био;н>гиче-ские ритмы. М., 1984. Т. 1. С. 54-69.
Баевский Р. М. Прогнозирование состояний на грани нормы и патологии. М., 1979. 295 с.
Балицкий К. П., Векслер И. Г., Винницкий В. Б. и др. Нервная система и противоопухолевая защита. Киев, 1983. 256 с.
Балицкий К. П., Векслер И. Г. Реактивность организма и химиотерапия опухолей. Киев, 1975. 253 с.
Барсукова Л. П., Гаркави Л. X., Котляревская Е. С, Марьяновская Г. Я.
Влияние регулирующих воздействий различных частотных характеристик
магнитного поля на развитие адаптационных реакций и циклические
изменения в митохонд-риях//Применение лазеров и магнитов в биологии и
медицине. Ростов н/Д, 1983. С. 23.
Барсукова Л. П., Котляревская Е. С, Марьяновская Г. Я. Фазные изменения
активности дегидро-геназ лимфоцитов при действии переменного магнитного
поля различных частот//Там же. С. 22.
Барсукова Л. П., Котляревская Е. С, Марьяновская Г. Я. К вопросу об
энергетическом гомеостазе организма при развитии различных
адаптационных реакций// Гомеостатика живых и технических систем.
Иркутск, 1987. С. 49—50.
Батрак Г. Е., Кудрин А. Н. Дозирование лекарственных средств экспериментальным животным. М., 1979. 168 с.
Бауэр Э. С. Теоретическая биология. М.; Л.* 1935. 208 с.
Бермант М. В., Гаркави Л. X., Захарова Н. П., Квакина Е. Б. и др.
Изменение некоторых иммуно-логических показателей при адаптационных
реакциях тренировки, активации и стресса//Регуляция иммунного
гомеостаза. Материалы докладов III Всесоюз. симпозиума. Л., 1982. С.
37—38.
Бирина Л. М., Гаркави Л. X., Квакина Е. Б., Кузьменко Т. С. и др.
Некоторые показатели иммуните-та в связи с характером адаптационных
реакций при химио-лучевом лечении онкологических больных//Актуальные
вопросы иммунологии и иммунопатологии в медицине и курортологии.
Пя-тигорск, 1987. С. 105—106.
Блехман И. И. Синхронизация динамических систем. М., 1971. 894 с.
Блехман И. И. Устойчивость движения. Аналитическая механика. Управление
движением. М., 1981. С. 55—66. Цит. по: Путилов А. А.
Системообразующая функция синхронизации в живой природе. Новосибирск,
1987. 144 с.
Бордюшков Ю. Н. О роли гипоталамо-гипофизарной системы в механизме
повышения резистент-ности организма (на модели опухолевого процесса):
Автореф. дис. ... докт. мед. наук. Ростов н/Д, 1971. 29 с.
Бордюшков Ю. Н. Состояние резистентности как результат многосвязного
регулирования со сто-роны функциональной системы адаптации//Адаптация и
проблемы общей патологии. Новоси-бирск, 1974. С. 68—70.
204
Бордюшков Ю. Н., Гашникова Л. И., Квакина Е. Б. Реакция эндокринной
системы на воздействия низкочастотным МП и ИМП//Материалы симпозиума
«Гигиеническая оценка магнитных полей». М., 1972. С.
48—52.
Браженко Н. А. Осложнения и осложненное течение туберкулеза органов
ды-хания/Сб. научных работ врачей ордена Ленина Ленинградского военного
округа. Ленинград, 1985. Ч. II. С. 72-74.
Брехман И. И. Женьшень. Л., 1957.
Брехман И. И. Элеутерококк. Л., 1968. 186 с.
Брехман И. И. Человек и
биологически активные вещества. Л.,
1976. 112 с.
Брехман И. И. Элеутерококк. М., 1984. 92 с.
Брюшков С. С. Коррекция естественной резистентности организма больного
пирогеналом как спо-соб предоперационной подготовки//Клиническая
хирургия. 1983. № 1. С. 42-44.
Бубнов В. Д., Кузьмина В. Е. Реакция щитовидной железы на действие
простых и комбинирован-ных стрессоров/Учен. зап. Пензенского пед.
ин-та. 1963. С. 51—69.
Бутов М. А. Значение адаптационных реакций организма для оценки влияния
Холодовых закали-вающих процедур. Теория и практика физической
культуры// Физкультура и спорт. 1984. № И. С. 30—32.
Василевский Н. И., Сороко С. И., Богословский М. М.
Психофизиологические аспекты адаптации человека в Антарктиде. Л., 1978.
208 с.
Васильев Н. В., Богинич Л, Ф. Влияние магнитных полей на процессы инфекции и иммунитета. Томск, 1973.
Васильев Н. В. Очерки о роли кроветворной ткани в антителообразовании. Томск, 1975. 300 с.
Васильев Н. В., Коляда Т. И. Неспецифические факторы иммунитета
человека на фоне адаптации к условиям Крайнего Севера//Вестник АМН
СССР. 1976. № 6. С. 66-70.
Введенский Н. Е. Физиологические явления с биологической точки зрения//
Физиология нервной системы. М., 1952. Т. 1. С. 566—574.
Венчиков А. И. Биотики. М., 1962.
Владимирский Б. М., Кисловский Л. Д. Космические воздействия и эволюция биосферы. М., 1986. 64 с.
Винер Н. Кибернетика, или управление и связь в животном и машине. 2-е изд. М., 1983. 294 с.
Виру А. А. Адренокортикальная активность при повторяющихся стрессориых
воздействи-ях//Вопросы эндокринологии. Тарту, 1974. С. 139—140.
Возрастная динамика сукцинатдегидрогеназной активности лимфоцитов у
физически активных и неактивных людей/'Комиссарова И. А., Лаврухина Г.
И., Гуд-кова Ю. В., Зивенко А. И. и др.//Регуляция энергетического
обмена и физиологическое состояние организма. М., 1978. С. 179
—183.
Волегов А. И. Устойчивость организма к злокачественным опухолям. М., 1987. 240 с.
Газенко О. Г., Генин А. М. Адаптация и стресс в космическом полете//
XII съезд Всесоюзного фи-зиологического общества им. И. П. Павлова.
Тбилиси, 1975. Т. 1. С. 143-144.
Гаркави Л. X. Влияние раздражений гипоталамуса на рост перевиваемых
сарком: Автореф. дис. ... канд. мед. наук. Ростов н/Д, 1962. 21 с.
Гаркави Л. X. Влияние раздражения (средней силы) гипоталамуса на рост
перевиваемых сар-ком//Вопросы клинической и экспериментальной
онкологии. Ростов н/Д, 1964. С. 30-31.
Гаркави Л. X. Влияние раздражений гипоталамуса «средней
силы» на рост некоторых эксперимен-тальных опухолей//Материалы IV
объединенной научной конференции медицинского и НИ ин-тов г. Ростова
н/Д. Ростов н/Д. 1968. С. 95— 96.
Гаркави Л. X. Значение силы раздражения гипоталамуса для получения
противоопухолевого эф-фекта//Бюл. экспер. биол. и мед. 1968. № 11. С.
95—96.
205
Гаркави Л. X. Об общей неспецифической адаптационной «реакции
активации», способствующей борьбе организма с опухолыо/'/Вопросы
клинической онкологии и нейроэндокринных нарушений при злокачественных
новообразованиях. Ростов н/Д, 1968. С. 341-348.
Гаркави Л. X. Адаптационная «реакция активации» и ее роль в
механизме противоопухолевого влияния раздражений гипоталамуса: Автореф.
дис. ... докт. мед. наук. Донецк, 1969. 30 с.
Гаркави Л. X., Гарибьян В. Р. Формула крови и вес вилочковой железы при
развитии общей неспе-цифической адаптационной «реакции
активации» у опухолевых и неопухолевых живот-ных//Клинические и
экспериментальные исследования в онкологии. Материалы Северо-Кавказской
онкологической конференции. Ростов н/Д, 1968. Ч. 1. С. 62-64.
Гаркави Л. X., Евстратова О. Ф., Коробейникова Е. П.
Морфофункциональ-ная характеристика органов тимико-лимфатической
системы при общих неспецифических адаптационных реакциях, вызванных
электростимуляцией эмоциогенных структур мозга//Функциональная
морфология лимфатических узлов и других органов иммунной системы и их
роль в иммунных процессах. М., 1983. С. 40—42. Гаркави Л. X.,
Квакина Е. Б. О принципе периодичности в реакции организма на магнитное
поле как неспецифический раздражитель//Влияние магнитных полей на
биологические объекты. Материалы III Всесоюз. симпозиума. Калининград,
1975. С. 18-19.
Гаркави Л. X., Квакина Е. Б. Адаптационные реакции организма и
многоуровневая регуляция го-меостаза//Физиологические и клинические
проблемы адаптации организма человека и животного к гипоксии,
гипертермии, гиподинамии и несяе-цифические средства восстановления.
М., 1978. С. 34—35.
Гаркави Л. X., Квакина Е. Б.
Диапазоны адаптационных реакций организ-
ма//Математическое моделирование биологических процессов. М., 1979. С. 27—33.
Гаркави Л. X., Квакина Е. Б. Адаптационные реакции организма, его резистент-
ность в связи с мышечной деятельностью//Проблемы физической культуры. Киев,
1982. С. 24-32.
Гаркави Л. X., Квакина Е. Б. Теория и практика адаптационных реакций
организма//Сравнительная эпидемиология
и диагностика основных локализаций
рака. М., 1984. С. 22-27.
Гаркави Л. X., Квакина Е. Б. О принципах использования
количественно-качественной закономер-ности развития адаптационных
реакций для повышения устойчивости к стрессу и борьбы с разви-вающимся
стрессом//Стресс, адаптация и функциональные нарушения. Кишинев, 1984.
С. 74.
Гаркави Л. X., Квакина Е. Б. О принципах лечебного действия магнитного
поля//Сравнительная эпидемиология и
диагностика основных локализаций рака.
С. 22-27.
Гаркави Л. X., Квакина Е. Б. Проявление закона единства и борьбы
противоположностей при раз-витии адаптационных
реакций//Философско~методический анализ противоречия как важнейшего
фактора в гомеостазе живых организмов и поведения малых коллективов.
Тезисы докл. Иркутск, 1985. С. 53—55.
Гаркави Л. X., Квакина Е. Б. Адаптационные реакции разных уровней
реактивности и взаимоот-ношения опухоли и организма//1У Всесоюз. съезд
онкологов. Тезисы докл. Л., 1986. С. 461—462.
Гаркави Л. X., Квакина Е. Б. О месте активационной терапии при этапном
лечении злокачествен-ных
новообразований//!!! Всероссийский
съезд онкологов.
Ростов н/Д, 1986. С. 558-560.
Гаркави Л. X., Квакина Е. Б. Представления о норме, предпатологии и
патологии в связи с систе-мой адаптационных реакций//Гомеостатика живых
и технических систем. Иркутск, 1987. С. 43—45.
Гаркави Л. X., Квакина Е. Б. Разные типы рез'истентности организма и роль
противоречия//Там же. С. 45—47.
Гаркави Л. X., Квакина Е. Б. О принципе диагностического лечения с целью
206
выявления групп риска и лечения заболеваний на начальных
стадиях//Онкологи ческяй компонент в диспансеризации населения. М.,
1987. С. 98—102.
Гаркави Л. X., Квакина Е. Б. Оценка и регуляция функционального
состояния с учетом адаптаци-онных реакций и состояний
ареактивности//Медицинские информационные системы. Междуве-домственный
тематический научный сборник. Таганрог, 1988. С. 64-71.
Гаркави Л. X., Квакина Е. Б., Барсукова Л. П., Котляревская Е. С. и др.
Характеристика сопряжен-ной дыхательной цепи печени крыс при
формировании адаптационных реакций//Митохондрии. Механизмы сопряжения и
регуляции. Пущино, 1981. С. 67-68.
Гаркави Л. X., Квакина Е. В., Барсукова Л. П., Котляревская Е. С. и др.
Фосфорилирующее дыха-ние гомогенатов печени крыс при периодическом
развитии реакций тренировки и стрес-са//Механизмы временной организации
клетки и их регуляции на различных уровнях. Тезисы докл. Пущино, 1983.
С. 51.
Гаркави Л. X., Квакина Е. Б., Барсукова Л. П., Шихлярова А. И. и др. О
некоторых закономерно-стях развития общих и местных неспецифических
адаптационных реакций//Вопросы клинической онкологии и нейроэндокринных
нарушений при злокачественных новообразованиях. Ростов н/Д, 1982. Вып.
5. С. 74—76.
Гаркави Л. X., Квакина Е. Б., Захарова Н. П., Шихлярова А. И. и др. К
вопросу о роли специфики и специализации при развитии общих
неспецифических адаптационных реакций организма//Там же. С. 74.
Гаркави Л. X., Квакина Е. Б., Котляревская Е. С, Барсукова Л. П. и др.
Сравнительная оценка влияния введения постоянной и меняющейся дозы
элеутерококка на некоторые показатели энерге-тического обмена в
нечени//Всесоюз. симпозиум. Молекулярные механизмы и регуляция
энерге-тического обмена. Тезисы докл. Пущино, 1986. С. 119-120.
Гаркави Л. X., Квакина Е. Б., Котляревская Е. С, Мамцева Л. И. и др.
Зависимость изменения ак-тивности дегидрогеназ лимфоцитов при старении
от развития стресса или реакции актива-ции//Труды IV Всесоюз. съезда
геронтологов и гериатров. Тезисы докл. Кишинев, 1982. С. 86—87.
Гаркави Л. X., Квакина Е. Б., Лупичев Л. Н. Связь неоднородности
временных шкал с уровнем ре-активности адаптационных реакций
организма//Примене-ние медицинской техники в хирургии. Тезисы докл.
Всесоюз. научной конференции. Иркутск, 1985. Ч. 1. С. 84-85.
Гаркави Л. X., Квакина Е. Б., Лупичев Л. Н., Файдыш Е. А. К динамике
конкурентного взаимодей-ствия при формировании триады адаптационных
реакций// Класс управления, использующий принцип противоречия, его
проявления в живых системах и возможности применения в технике. Тезисы
к научной конференции. Иркутск, 1984. С. 40-42.
Гаркави Л. X., Квакина Е. В., Лупичев Л. Н., Файдыш Е. А. К возможности
возникновения негаэн-тропийного эффекта при отдельных режимах конфликта
в системе физиологического гомеоста-за//Философско-методологический
анализ противо речия как важнейшего фактора в гомеостазе живых
организмов и поведении малых коллективов. Тезисы докл. С. 55—57.
Гаркави Л. X., Квакина Е. Б., Мулатова А. К., Евстратова О. Ф. и др.
Морфологическое исследо-вание спинного мозга и периферической нервной
системы крыс при общих неспецифических адаптационных реакциях//Изв.
СКНЦ ВШ. № 1212. 1984. С. 84.
Гаркави Л. X., Квакина Е. Б., Мулатова А. К., Тарасьянц Ф. М. О
применении в эксперименте про-тивоопухолевых препаратов в малых
дозах//Вопросы онкологии. 1981. Т. 27. № 7. С. 61-66.
Гаркави Л. X., Квакина Е. Б., Мулатова А. К., Шихлярова А. И. О
повышении противоопухолевой резистентности с помощью малых доз
адреналина//Там же. 1980. Т. 26. № 6. С. 48-53.
Гаркави Л. X., Квакина Е. В., Серегин Ю. В., Файдыш Е. А. О
многомерности силовых отношений в системе биологического
гомеостаза//Применение медицинской
207
техники в хирургии. Тезисы докл. Всесоюз. научной конференции. Ч. 1. С.
81—83. Гаркави Л. X., Квакина Е. В., Уколова М. А. Адаптационная
«реакция активации» как средство воздействия на гипоталамус
с целью торможения преждевременного старения//Физиологическое понятие
возрас-тной нормы. Тезисы Всесоюз. симпозиума с участием иностранных
ученых. Л. 1971. С. 45—46.
Гаркави Л. X., Квакина Е. Б., Уколова М. А. Роль неспецифических
адаптационных реакций (стрес-са и активации) во взаимоотношениях
опухоли и орга-низма//Опухоль и организм. Материалы на-учной
конференции. Киев, 1973. С. 81-82.
Гаркави Л. X., Квакина Е. Б., Уколова М. А. О возможности управления
противоопухолевой рези-стентностыо организма//Воиросы клинической
онкологии и пейроэндокринных нарушений при злокачественных
новообразованиях. Вып. 3. С 216—220.
Гаркави Л. X., Квакина Е. Б., Уколова М. А. Изменение состава белой
крови как критерий адапта-ционных реакций организма при воздействии
магнитными полями и другими неспецифическими агентами//Магнитное поле в
медицине. Материалы к симпозиуму «Влияние искусственных полей на
биологические объекты». Фрунзе. 1974. Т. 100. С. 23-25.
Гаркави Л. X., Квакина Е. Б., Уколова М. А. Роль адаптационных реакций
в патологических про-цессах и простые критерии этих реакций у
людей//Регуляция энергетического обмена и устойчи-вость
организма. Пущино, 1975. С. ,172—182.
Гаркави Л. X., Квакина Е. Б., Уколова М. А. Геронтологические аспекты
изучения неспецифиче-ской адаптационной реакции активации//Тезисы XII
съезда Всесоюз. физиологического общества им. И. И. Павлова. Тбилиси,
1975. Т. 3. С. 94—95. Гаркави Л. X., Квакинй Е. Б., Уколова М. А.
Адаптационные реакции организма и противоопухолевая
резистентность//Тезисы докл. II Всесо-юз. съезда патофизиологов.
Ташкент, 1976. Т. 2. С. 149-150. .
Гаркави Л. X., Квакина Е. Б., Уколова М. А. Адаптационные реакции
тренировки и активации как средство преодоления хронического стресса
при старении// III Всесоюзный съезд геронтологов и гериатров. Киев,
1976. С. 71.
Гаркави Л. X,, Квакина Е. Б., Уколова М. А. Исследование адаптационных
реакций тренировки и активации для неспецифической
терапии//Физиологические и клинические проблемы адаптации организма
человека и животного к гипоксии, гипертермии, гиподинамии и
неспецифические сред-ства восстановления. С. 240— 241.
Гаркави Л. X., Квакина Е. Б., Уколова М. А. Магнитные поля,
адаптационные реакции и резнстент-ность организма//Реакции
биологических систем на магнитные поля. М„ 1978. С. 131-148.
Гаркави Л. X., Квакина Е. Б., Уколова М. А. Адаптационные реакции и рези-стентность организма. Ростов н/Д, 1979. 119 с.
Гаркави Л. X., Квакина Е. Б., Уколова М. А. К вопросу о теоретическом
обосновании магнитотера-пии в онкологии//Ш Всероссийский съезд
онкологов. С.
574-576.
-.
Гаркави Л. X., Квакина Е. Б., Уколова М. А., Захарюта Ф. М. и др.
Повышение сопротивляемости организма с помощью адаптационных реакций
тренировки и активации на разных уровнях реак-тивности организма
(активационная терапия)// Методические рекомендации. Ростов н/Д, 1982.
12 с.
Гаркави Л. X., Квакина Е. Б., Уколова М. А., Захарова Н. П. и др. О
противоопухолевом действии магнитных полей в эксперименте//Применение
лазеров в биологии и медицине. Тезисы докл. Рос-тов н/Д, 1983. С. 34.
Гаркави Л. X., Квакина Е. Б., Уколова М. А., Мулатова А. К. и др. О
морфо-функциональных изме-нениях в тимико-лимфатической системе при
развитии общих неспецифических адаптационных реакций тренировки и
активации//Регуляция иммунного гомеостаза. Материалы докладов III
Все-союз. симпозиума. Л., 1982. С. 43-44.
Гаркави Л. X., Квакина Е. Б., Шейко Е. А. Люминесцентные и морфометри-
208
ческие параметры клеток щитовидной железы крыс при введении различных
доз адреналина и иммобилизационном стрессе//Проблрмы эндокринологии М .
1987 С. 45-47.
Гаркави Л. X., Квакина Е. Б., Уколова М. А., Шихлярова А. И. и др.
Изменения адаптационных ре-акций организма и его резиетентности под
влиянием магнитных полей//Электромагнитные ноля в биосфере.
М., 1984. Т. 2. С. 46—56.
Гаркави Л. X., Квакина Е. Б., Шихлярова А. И., Тарасъянц Ф. М.
Количественно-качественная за-кономерность биологического действия
электромагнитных полей: нелинейный характер действия ПеМП на уровне
организма н ткани//Био-логическое действие электромагнитных полей.
Пущино, 1982. С. 60 — 61.
Гаркави Л. X., Колесникова В. А., Андреева Л. А. Влияние оливомицина,
облучения, адреналина и их сочетаний на саркому-45//Вопросы клинической
онкологии и нейроэндокринных нарушений при злокачественных
новообразованиях. Вып. 2. С. 37-40.
Гаркави Л. X., Мацанов А. К. Изменение периферической крови и
морфологии коры надпочечни-ков под влиянием различных доз
облучения/'/Функцнональное состояние желез внутренней секре-ции при
опухолевом процессе. Ростов н/Д. 1973. С. 182-186.
Гаркави Л. X., Рогозная А. В. Изменения системы аммиак — глютамин
— глютаминовая кислота мозга при раздражениях гипоталамуса,
влияющих на рост перевивных сарком//Материалы конфе-ренции но
опосредованному воздействию на опухолевый процесс. Л., 1963. С. 34-35.
Гаркави Л. X., Старкова А. А., Якунина Н. Н. Содержание нуклеиновых
кислот в некоторых тка-нях при развитии первой фазы общей
неспецифической реакции активации//Материалы Северо-Кавказской
онкологической конференции. Ростов н/Д, 1968. Ч. 1. С. 58-59.
Гаркави Л. X., Трапезонцева Р. А. Об изменениях нуклеиновых кислот в
мозгу и тканях внутрен-них органов при изменении функционального
состояния гипоталамуса длительным электрическим раздражением//Х
Всесоюз. съезд физиологического общества им. И. 11. Павлова.
М., 1964. Т. 2. С. 191.
Гаркави Л. X., Уколова М. А., Григорьева Е. Г. Изменение факторов
свертывающей и антисверты-вающей систем крови в первую фазу различных
адаптационных реакций//Вопросы клинической онкологии и нейроэндокринных
нарушений при злокачественных новообразованиях. Выи. 2. С. 41—43.
Гаркави Л. X., Уколова М. А., Квакина- Е. В. Развитие реакции активации
как способ комплексного повышения функциональной активности эндокринных
желез при опухолевом ироцес-се//Функциональное состояние желез
внутренней секреции при опухолевом процессе. Ростов н/Д, 1973. С.
34—39.
Гаркави Л. X., Уколова М. А., Квакина Е. Б. Закономерность развития
качественно отличающихся общих неспецифических адаптационных реакций
организма. Диплом на открытие № 158 Комите-та Совета Министров СССР по
делам изобретений и открытий//Опфытия в СССР. М.. 1.975. С. 56 —
(П.
Гашников а Л. И., Бордюшков Ю. И., Квакина, Е. Б. Об изменении функции
щитовидной железы под влиянием магнитных нолей//Рсакция биологических
систем на слабые магнитные поля. Мате-риалы Всесоюз. симпозиума. М.,
1971 С 135— 137.
Генкин А. А. Статистический анализ ЭЭГ как общая проблема анализа
колебательных процессов, протекающих в физиологических
системах//Математнчсский анализ электрических явлений го-ловного мозга.
М., 1965. С. 72—86.
Гервазиева И. Д. Изменение функционального состояния коры надпочечников
в условиях «на-пряжения»/Учен. заи.
Черновицкого мед. ин-та. 1963. № 40. С. 45.
Глансдорф П., Пригожим И. Термодинамическая теория структуры, устойчивости и флуктуации. М., 1973. 280 с.
Глухман М. В., Ступин Г. К. Адаптационные реакции как метод определения
функционального состояния спортсменов//!! Всероссийский съезд по
врачебному контролю и лечебной физкультуре. Ярославль, 1979. С. 154
— 155.
209
Голубев Г. Ш. Индивидуальная реактивность при травматической болезни
(особенности течения, прогноз и диагностика осложнений): Автореф. дис.
... канд. мед. наук. Ростов н/Д, 1985. 24 с.
Голубев Г. Ш., Локшина Е. Г., Румбешт Л. М., Киселева Л. В. Особенности
иммунологической ре-активности у больных с переломами длинных трубчатых
костей в посттравматическом перио-де//УП Северо-Кавказская конференция
по иммунологии, аллергологии и
молекулярной био-логии. Краснодар,
1983. С. 105—106.
Горизонтов П. Д., Протасова Т. Н. Роль АКТГ и кортикостероидов в патологии. М., 1968. 334 с.
Городилова В. В. Клинические аспекты иммунологии
опухолей//Теоретические и клинические ас-пекты иммунологии опухолей.
Кемерово, 1975. С. 9 — 59.
Гращенков Н. И. Гипоталамус и его роль в физиологии и патологии. М., 1965. С. 10.
Гращенков Н. И. Гипоталамус. Физиология и патофизиология гипоталамуса//
Докл. 1-й Всесоюз. конференции по физиологии и патологии гипоталамуса.
М., 1969. С. 9.
Григорьева Е. Г. О состоянии свертывающей системы крови при развитии
неспецифических адап-тационных реакций у опухолевых и неопухолевых
животных// Система свертывания крови и фиб-ринолиз. Тезисы IV Всесоюз.
конференции. Саратов, 1975. С. 363-364.
Гудвин В. Временная организация клетки: Динамическая теория внутриклеточных регуляторных процессов. М., 1966. 251 с.
Гудвин Б. Аналитическая физиология клеток и развивающихся организмов. М., 1979. 287 с.
Данишевский Г. М. Патология человека и профилактика заболеваний на Севере. М., 1968. 412 с.
Дардымов И. В. Женьшень, элеутерококк. М., 1976. 189 с.
Деряпа Н. Р., Рябинин И. Ф. Адаптация человека в полярных районах. Л., 1977. 296 с.
Дильман В. М. Подходы к увеличению продолжительности жизни человека в
свете элевационного механизма развития, старения и формирования
специфической возрастной патоло-гии//Геронтология и гериатрия. 1975;
Биологические возможности увеличения продолжительно-сти жизни. Киев,
1976. С. 129—137.
Дильман В. М. Мутационно-метаболическая модель возникновения рака и
прогрессии опухолево-го процесса//Вопросы
онкологии. 1976. Т. 22.
№ 8. С. 3—16.
Дильман В. М. Эндокринологическая онкология. Л., 1983. 407 с.
Дильман В. М. Четыре модели медицины. Л., 1987. 286 с.
Дубров А. Л. Влияние гелиогеофизических факторов на проницаемость
мембран и суточную рит-мичность выделения органических веществ корнями
растений// Докл. АН СССР. 1969. Т. 187. С. 14-29.
Дубров А. П. Симметрия биоритмов и реактивности. М., 1987. 175 с.
Дэвис Ф. Онтогенез циркадианных ритмов//Биологические ритмы. М., 1984. Т. 1. С. 292-309.
Еремина С. А. О некоторых механизмах стресса//Стресс и его
патогенетические механизмы. Мате-риалы Всесоюз. симпозиума. Кишинев,
1973. С. 22—23.
Еремина С. А., Бойченко А. Е., Белякова Е. И., Рогачева Е. А.
Инициальный период стресса и его роль в адаптации//Стресс, адаптация и
функциональные нарушения. С. 85—86.
Ерохин В. Г Методологические и социокультурные предпосылки построения
общемедицинской теории: Автореф. дис. ... докт. филос. наук. Ростов
н/Д, 1985. 47 с.
Жаботинский А. М. Колебательные химические реакции в гомогенной среде и
смежные пробле-мы//Колебательные процессы в биологических и химических
системах. Пущино. 1967. Т 2. С. 288.
Жуков Б. Н. Научное обоснование применения магнитных полей в медицине//
Биологические эффекты
электромагнитных полей: вопросы
их использования и
210
нормирования. Пущино, 1986. С. 108—122.
Жуков Б. Н., Труфанов Л. А., Мусиенко С. М. К оценке
окислительно-восстановительных процес-сов в биологических объектах при
воздействии магнитного поля//Магнитобиология и
магнито-терапия в медицине. Витебск,
1980. С. 51 — 52.
Западнюк В. И. Фармакотерапевтическая эффективность и механизмы
действия некоторых гери-атрических препаратов//Геронтология и
гериатрия. 1975; Биологические возможности увеличения продолжительности
жизни. С. 180 —188.
Западнюк В. И. Лекарственные средства, ослабляющие явление
преждевременного старе-ния//Вестник АМН СССР. 1984. № 3. С. 65 —
70.
Зарецкая Ю. М. Клиническая пммуногенетика. М., 1983. 206 с.
Захарова Н. П., Бермант М. В., Самохвалова Л. И., Дерибас В, И.
Некоторые показатели характера адаптационных реакций на разных уровнях
реактивности при активационной терапии//Вопросы клинической онкологии и
нейроэндокринных нарушений при злокачественных новообразованиях. С. 74.
Захарова Н. П., Гаркави Л. X., Квакина Е. Б., Лупичев Л. Н. и др.
Активацион-ная терапия неспе-цифических воспалительных процессов
женской половой сферы с помощью магнитопункту-ры//Современные проблемы
рефлексодиагностики и рефлексотерапии. Ростов н/Д, 1984. С. 151 —
153.
Захарова Н. П., Квакина Е. Б. Активационная магнитотерапия кардионевро
зов//Применение лазе-ров и магнитов в биологии и медицине. Тезисы докл.
С. 58.
Захарюта Ф. М. Защитное влияние адаптационных реакций тренировки и
активации, вызываемых магнитным полем, при ионизирующем облучении
крыс//Там же. С. 118.
Захарюта Ф. М. Возможность увеличения действующей дозы
противоопухолевых препаратов с помощью магнитных полей//Магнитные поля
в биологии, медицине и сельском хозяйстве. Тезисы докл.
научно-практической конференции. М., 1985. С. 71-72.
Захарюта Ф. М. К механизму защитного влияния магнитного поля малой
интенсивности при хи-миотерапии опухолей в эксперименте//Механизмы
биологического действия электромагнитных излучений. Цущино, 1987. С.
173.
Захарюта Ф. М. Модификация химиотерапии опухолей путем регуляции
неспецифической рези-стентности организма: Автореф. дис. ... канд.
биол. наук. Киев, 1989. 26 с.
Кавецкий Р. Е. Взаимодействие организма и опухоли. Киев, 1977. 236 с.
Казначеев В. П. Современные аспекты
адаптации. Новосибирск, 1980.
198 с.
Казначеев В. П. Учение о биосфере. М., 1985. 77 с.
Казначеев В. П., Баевский Р. М. Индивидуальные особенности
адаптационных реакций у человека и проблема донозологической
диагностики//Адаптация и проблема общей патологии. Новоси-бирск, 1974.
Т. 2. С. 9—13.
Казначеев В. П., Казначеев С. В. Адаптация и конституция человека. Новосибирск, 1986. 119 с.
Казначеев В. П., Куликов В. Ю. Некоторые особенности патологии человека
на Крайнем Севе-ре//Механизмы адаптации человека в условиях высоких
широт. Л., 1980. С. 115-180.
Казначеев В. П., Михайлова Л. П. Биоинформационная функция естественных электромагнитных полей. Новосибирск, 1985. 181 с.
Календо Г. С. О возможности адаптационного синдрома стресса на
клеточном уровне и его роли в реакции клетки на облучение//Успехи
современной биологии. 1972. Т. 73. № 1. С. 53.
Календо Г. С. Ранние реакции клеток на ионизирующее излучение и их роль в защите и сенсиби-лизации. М., 1982. 96 с.
Календо Г. С, Винская Н. П. Надежность систем защиты и адаптационный
комплекс реакций на уровне клеточной популяции//Системы надежности
клетки. Киев, 1977. С. 104-118.
Караш Ю. М., Балина Ю. Д., Кононова Э. Ф., Чижов А. Я. и др. Об
адаптационных реакциях боль-ных с хроническим неспецифическим
воспалением придат-
211
ков матки//Акушерство и гинекология. 1984. № 9. С. 61—64.
Караш Ю. М., Стрелков Р. Б., Чижов А. Я. Нормобарическая гипоксия в лече-нии, профилактике и реабилитации. М., 1988. 351 с.
Кассиль Г. Н. Внутренняя среда организма. М., 1983. 224 с.
Кассиль Г. Я., Матлина Э. Ш. Симпатоадреналовая система при стрессе//Стресс и его патогенети-ческие механизмы. С. 24—26.
Кацман М. В., Сидоренко Ю. С, Айзенштарк Э. А. Здоровье без лекарств. Ростов н/Д, 1986.
Квакина Е. Б. Повышение неспецифической противоопухолевой
резистентности организма с по-мощью бесконтактного раздражения
гипоталамуса: Автореф. дис. .. докт. биол. наук. М., 1972. 41 с.
Квакина Е. Б. Адаптационные реакции разных уровней реактивности как
не-специфическая основа лечебного действия магнитного поля//Магнитные
поля в био-иогии, медицине и сельском хозяй-стве. С. 24—26.
Квакина Е. Б., Барсукова Л. П., Шихлярова А. //.'Влияние ПеМП малой
интенсивности и различной частоты на лимфоидные органы//Функциональная
мор-рология лимфатических узлов и других органов иммунной системы и их
роль в иммунных процессах. Тезисы докл. Всесоюз. научной конференции.
М., 1983. С. 40.
Квакина Е. Б., Гаркави Л. X. Принцип периодичности в реакции организма
на магнитные поля на-растающей интенсивности//Физико-математические и
биологические проблемы действия элек-тромагнитных нолей и ионизации
воздуха. М., 1975. С. 52-54.
Квакина Е. В., Гаркави Л. X. О некоторых принципах организации
гомео-стэза//Примененне меди-цинской техники в хирургии. Тезисы докл.
Всесоюз. научной конференции. Ч. 1. С. 83—84.
Квакина Е. Б., Гаркави Л. X., Уколова М. А. Управление резистентностью
организма на основе адаптационных реакций. Таганрог,
1984. С. 46 — 56.
Квакина Е. Б., Гаркави Л. X., Уколова М. А., Тарасьянц Ф. М. и др.
Общие адаптационные реакции тренировки и активации и защита организма
от повреждающего действия противоопухолевых препаратов//Актуальные
проблемы экспериментальной химиотерапии опухолей. Черноголовка, 1980.
Т. 2. С. 159—162.
Квакина Е. Б., Григорьева Е. Г., Марьяновская Г. Я. Изменения процесса
свертывания крови под влиянием магнитного поля, способного вызывать
противоопухолевый эффект//Вопросы клиниче-ской онкологии и
нейроэндокринных нарушений при злокачественных новообразованиях. Вып.
2. С. 91—93.
Квакина Е. Б., Додис Л. И., Куликова Н. И., Богуславская 3. К. и др.
Адаптационные реакции орга-низма на санаторном этапе реабилитации
больных с инфарктом миокарда//Ранняя диагностика и профилактика
сердечно-сосудистых заболеваний. Новосибирск, 1983. Ч. II. С. 158-159.
Квакина Е. Б., Квакин С. Д., Котляревская Е. С. Исследование
возбудимости гипоталамуса и ее изменение под влиянием магнитных
полей//Материалы XVII научной конференции физиологов юга РСФСР.
Ставрополь, 1969. С. 137 — 139.
Квакина Е. Б., Котляревская Е. С, Квакин С. Д. Изменение возбудимости
гипоталамуса под влия-нием магнитного поля//Материалы II Всесоюз.
совещания по изучению влияния магнитных полей на биологические объекты.
М., 1969. С. 103— 106.
Квакина Е. Б.,. Николаева В. И., Уколова М. А. О характере
неспецифической реакции организма в ответ на длительное воздействие
низкочастотным переменным магнитным полем//Реакция биоло-гических
систем на слабые магнитные поля. Материалы Всесоюз. симпозиума. М.,
1971. С. 122—124.
Квакина Е. В., Рудой Ф. М. О роли сульфгидрильных групп в
неспецифической противоопухоле-вой резистентности//Вопросы клинической
онкологии и нейроэндокринных нарушений
при злокачественных новообразованиях.
С. 229—233.
Квакина Е. Б., Уколова М. А. О различных адаптационных реакциях в
зависимости от силы воз-действия магнитного поля//Материалы II
Всесоюз. совещания
212
по изучению влияния магнитных полей
на биологические объекты. С.
107 — 110.
Квакина Е. Б., Уколова М. А., Гаркави Л. X., Арефьева Л. М. и др.
Изменение активности холинэ-стеразы при росте опухолей и
противоопухолевых воздействиях на центральную нервную систе-му//Вопросы
клинической онкологии и нейроэндокринных нарушений при злокачественных
ново-образованиях. С. 94—97.
Квакина Е. В., Шибкова С. А., Исаджанова С. X. О морфологических
изменениях в гипоталамусе при рассасывании опухолей под влиянием
переменного магнитного поля//Вопросы онкологии. 1974. Т. 20. С.
89—92.
Кикут Р. П. Влияние магнитных полей на систему крови и кровообращение//
Реакции биологиче-ских систем на магнитные поля. М., 1978. С.
149—166.
Кикут Р. П., Лиепа М. Э., Витолс Э. А. и др. Возможности применения
магнитных полей в нейро-хирургии//Биологические эффекты
электромагнитных полей. Вопросы их использования и норми-рования.
Пущине, 1986. С. 123 — 129.
Кикут Р. /7,, Лиепа М. Э., Крцминя Г. А., Кикуте С. Р. др.
Возможности применения магнитобио-логических эффектов в
нейрохирургии//Вопросы курортологии, физиотерапии и лечебной
физиче-ской культуры. М., 1981. № 4. С. 18 — 24.
Кириллов О. А. Изучение некоторых механизмов адаптационного действия
элеутерококка: Авто-реф. дис. ... канд. биол. наук. Томск, 1964.
Кисловский Л. Д. Реакции живых систем на слабые адекватные им
воздей-ствия/'/Электромапштные поля в биосфере. М., 1984. Т. 2. С.
6—16.
Клетке Г. Э. Адаптационные реакции организма у хирургического
контингента пульмонологиче-ских больных: Автореф. дис. ... канд. мед.
наук. Челябинск, 1990. 21 с.
Клетке Г. Э., Котляров С. В., Бехтерева Е. И. Адаптационные реакции
организма и их изменение при хирургическом лечении рака
легкого//Клиническая хирургия. 1984. № 5. С. 17-18.
Клецкин С. 3. Хирургический стресс и регуляция физиологических функций. М., 1983. С. 85.
Коган А. Б. Методика хронического вживления электродов для отведения потенциалов и раздра-жения мозга. М., 1952.
Коган А. Б. Рефлекторная деятельность нервной системы
позвоночных//Физиология человека и животных/Под ред. А.
Б. Когана. М., 1984. Ч.
II. С. 121 — 169.
Коломиевский М. Л. Адаптационные реакции у больных ишемической болезнью
серд-ца//Клиническая медицина. 1982. № 7. С. 32 — 35.
Комфорт А. Биология старения. М., 1967.
Комурджисв X. А., Квакина Е. Б., Савина С. А. и др. К вопросу об
особенностях радиобиологиче-ских реакций в постоянном магнитном
поле//Материалы XXIV научной сессии Московского НИ онкологического
института им. Герцена, Ростовского НИ онкологического института и
Сочинско-го городского онкологического диспансера. М., 1967. С.
334—339.
Кондрашова М. Н. Регуляция янтарном кислотой энергетического
обеспечения и функционально-го состояния ткани: Автореф. дис. ... докт.
биол. наук. Пущи-ио, 1971. 24 с.
Кондрашова М. Н. Участие митохондрий в развитии адаптационного синдрома. Пущ шш, 1974.
Кондрашова М. Н. Основные понятия биоэнергетики, используемые в
функциональных исследо-ваниях. Подвижность метаболических реакций
митохондрий//Ре-гуляция энергетического обмена и устойчивость
организма. Пущино, 1975. С. 67 — 82.
Кондрашова М. Н. Живое состояние с позиций
биоэнергетнки//Методологиче-ские и теоретиче-ские проблемы биофизики.
М., 1979. С. 200—212.
Кондрашова М. Н., Григоренко Е. В., Бабский А. М., Хазанов В. А.
Гомео-стазирование физиологи-ческих функций на уровне
мнтсхондрий//Молекулярньге механизмы клеточного гомеостаза.
Но-восибирск, 1987. С. 40—66.
Корнева Е. А., Клименко В. М., Шхинек Э. К. Нейрогуморальное обеспечение иммунного гомеоста-за. Л., 1978. 176 с.
213
Коробейникова Е. П. Изменения а-ритма человека при общих
неспецифических адаптационных реакциях, вызванных ПеМП//Применение
лазеров и магнитов в биологии и медицине. Тезисы докл. С. 64.
Косицкий Г.
И
-В. М. Нервная система и стресс.
М., 1970. 200 с.
Котляревская Е. С. Исследование функционального состояния
гипоталамиче-ской области голов-ного мозга при противоопухолевом
действии магнитных полей: Автореф. дис. ... канд. биол. наук. Ростов
н/Д, 1974. 24
с.
Котляревская Е. С, Маръяновская Г. Я., Барсукова Л. П., Коробейникова
Е. П. Активность дегид-рогеназ лимфоцитов периферической крови у крыс
при развитии адаптационных реакций трени-ровки, активации и
стресса//Вопросы клинической онкологии и нейроэндокринных нарушений при
злокачественных новообразованиях. С. 79-83.
Кравков Н. Я. О пределах чувствительности живой протоплазмы//Успехи современной биологии. 1924. № 3, 4.
Красногорская Н. В., Сперанский А. П., Десницкая М. М. Влияние внешних
факторов на организм и их роль в медикаментозном
лечении//Электромагнитные поля в биосфере. Биологическое дейст-вие
электромагнитных полей. М. 1984. Т. II. С. 249-256.
Крыжановский Г. И. Биоритмы и закон структурно-функциональной временной
дискретности биологических процессов//Биологические ритмы в механизмах
компенсации нарушенных функ-ций. М., 1973. С. 20—34.
Крыжановский Г. Н. Детерминантные структуры в патологии нервной системы. М., 1980. 360 с.
Крыжановский Г. Н. И лечение должно быть комплексным, системным//Бу-дущее науки. 1987. Вып. 20. С. 133-147.
Кудрин А. Я. Фармакология с основами патофизиологии. М., 1977. 550 с.
Кулик Г. И. Изменение количества сульфгидрильных групп в сыворотке
крови и ткани перевивае-мых опухолей крыс под влиянием алкилируюших
противоопухолевых препаратов//Вопросы онко-логии. 1964. Т. 10. № 7. С.
70—74.
Курышёв И. Е. Биоритмические изменения мембраны клеток при действии
цАМФ и цГМФ в плане поддержания гомеостаза//Гомеостатика живых и
технических систем. С. 51.
Лазарев Я. В. Сопоставление неспецифических защитных реакций, влияющих
на генерализацию инфекций и метастазирование опухолей//Вопросы
онкологии. 1962. Т. 8. № П..С 20-28.
Лазарев В. В. Адаптогены и рак//Материалы конференции по
опосредованному воздействию на опухолевый процесс. Л., 1963. С.
52—55.
Лазарев Н. В. Фармакологические методы в профилактике
рака//Антибласто-могенная терапия предраковых заболеваний. Материалы
конференции Казахского НИИ онкологии и радиологии. Алма-Ата, 1968. С.
117 — 121.
Макеев В. Б. Экспериментальное исследование физиологического действия
электромагнитных по-лей инфранизкой частоты на систему крови животных:
Автореф. дис. ... канд. биол. наук. Симфе-рополь, 1979. 25 ,с.
Макеева Е. Я. Система интенсивной активизирующей психотерапии затяжных
неврозов в специа-лизированном стационаре: Автореф. дис. ... канд.
биол. наук. М., 1981. 17 с
Макеев В. Б., Темурьянц Н. А. Исследование частотной зависимости
биологической эффективно-сти магнитного поля в диапазоне микропульсаций
геомагнитного ноля (0,01 — 100 Гц)//Проблемы космической
биологии. Влияние солнечной активности на биосферу. М., 1982. Т. 43. С.
116—128..
Макеев В. В., Темурьянц Н. А., Владимирский Б. М., Тишкина О. Г.
Физиологически активные ин-франизкочастотные магнитные
поля//Электромагнитные поля в биосфере. М., 1984. Т. 2. С. 62-72, 57-62.
Маньковский Н. Б., Минц А. Я., Литовченко С. В., Белоног Р. П. Динамика
функционального со-стояния центральной нервной системы у лиц пожилого
возраста под влиянием активного двига-тельного режима//Двигательная
активность и старение. Киев, 1969. С. 285-293.
214
Маньковский Н. Б., Минц А. Я., Белоног Р. П. К характеристике
возрастных изменений нервной системы человека при
старении//Физнологическое понятие возрастной нормы. Тезисы Всесоюз.
симпозиума с участием иностранных ученых. С. 26—28.
Маръяновская Г. Я. О механизме центрального и периферического действия
магнитных полей (на модели опухолевого процесса): Автореф. дис. ...
канд. биол. наук. Ростов н/Д, 1974. 24 с.
Марьяновская Г. Я., Барсукова Л. Я., Котляревская Е. С. и др. Изменения
некоторых показателей энергетического обмена при росте и регрессии
опухоли//Все-союзный семинар «Молекулярные механизмы и регуляция
энергетического обмена». Тезисы докл. Пущино, 1986. С. 136.
Марчук Г. И. Математические модели в иммунологии. М., 1980.
Меделяновский А. М. Энергетиче-ские аспекты гомеостаза//Гомеостазы и
гомео-статные сети управления, их
приложения в биологических
природных и технических системах. Иркутск, 1981. С.
41—42.
Меделяновский А. М.,
Гуськов С, В., Килина
Т. С. и др.
Количественные критерии оценки
состояний функциональной системы,
обеспечивающей газовый гомеостаз//Вестник Академии
медицинских наук СССР. М., 1985. № 2. С.
63—70. Меерсон Ф. 3. Основные
зако-номерности индивидуальной адаптации/Физиология
адаптационных процессов (Руководство по физиологии). М., 1986. С.
10—71. Меерсон Ф. 3. Общий механизм адаптации и роль в нем
стресс-реакции, основные стадии процесса//Там же. С. 77 — 123.
Меерсон Ф. 3. Адаптация к стрессорным ситуациям и стресс-лимитирующие системы организ-ма//Там же. С. 521—631.
Меерсон Ф. 3., Пшенникова М. Г. Адаптация к стрессорным ситуациям и физическим нагрузкам. М., 1988. 253 с.
Механизмы адаптации человека в условиях высоких шрот/Под ред В. П. Каз-начеева. М., 1980. 197 с.
Митюшов М. И. Гипофизарно-адреналовая система и стресс//Гипофизарио-ад^
реналовая система и мозг. Л. 1976. С. 192 — 204.
Моисеева Н. И., Сысуев В. М. Временная среда и биологические ритмы. Л., 1981. 126 с.
Мороз Ю. А., Никитин В. П., Ставицкий Л. И. Молекулярные и гормональные
сдвиги при экспе-риментальном продлении жизни//Успехи физиологических
наук. 1976. Т. 7. № 2. С. 116-129.
Музалевская И. И. Характеристика возмущенного геомагнитного поля как раз дражите-ля//Проблемы космической биологии. М., 1973.
Музалевская Н. И., Шушков Г. Д. Исследование биотронности параметров
слабого магнитного по-ля диапазона сверхнизких частот//Реакции
биологических систем на магнитные поля. С. 199—208.
Мулатова А. К. Морфофункциональная характеристика лимфоидных органов
крыс при адаптаци-онных реакциях//Функциональная морфология
лимфатических узлов и других органов иммунной системы и их роль в
иммунных процессах. Тезисы докл. Всесоюз. научной конференции. С. 120.
Мулатова А. К., Уколова М. А., Тарасъянц Ф. М., Евстратова О. Ф.
Защитное влияние магнитных полей по отношению к тимико-лимфатической
системе при химиотерапии опухолей//Актуальные вопросы магнитобиологии и
магнитотерапии. Ижевск, 1981. С. 47.
Муравов И. В. Двигательная активность, биологический возраст и продолжительность жиз-ни//Геронтология и гериатрия. 1975.
Нарциссов Р. П., КомиссароваЛ. А. Энзиматический статус лейкоцитов в
оценке индивидуального развития//Физиологическое понятие возрастной
нормы. Тезисы Всесоюз. симпозиума с участием иностранных ученых. С.
38—40.
Насонов Д. И., Александров В. Я. Реакция живого вещества на внешние воз
действия (денатурационная теория повреждения и раздражения). М., 1940. 420 с.
Нахилъницкая 3. Я. Особенности реакции различных функциональных систем
215
но воздействие сильных магнитных полей
и возможности адаптации//Биологичс-скоо дейст-вие
электромагнитных нолей. Пущино, 1982. С, 89 — 90.
Нечуткий А. И. Информационные аспекты гомеостазирующей функции
акупунктурных сис-тем//Современные проблемы рефлексотерапии. Тезисы
докл. Все-союз, конференции по рефлек-сотерапии. Волгоград, 1987. С.
30—32.
Нечуткий А. И., Николаев Н. А. Физиологические принципы контроля
функций организма (адап-тивного управления)//Известия АН Лат. ССР.
Рига, 1986. 7 (468). С. 37—46.
Никитин Н. В. Периодическое сдерживающее рост питание и пролонгирование
жиз-ни//Геронтология и гериатрия. 1975; Биологические возможности
увеличения продолжительности жизни. С. 73—77.
Николаев Л. А. Основы физической химии биологических процессов. 2-е изд. М., 1976. 260 с, Цит. по: Путилов А. А. Указ. соч.
Николаев Л. А.//Термодинамика и регуляция биологических процессов. М.,
1984. С. 255—259. Цит. по: Путилов А. А. Указ.
соч.
Ногаллер А. М., Бутов М. А., Калыгина Т. А. Иммунологическая
резистент-ность и адаптационные реакции при закаливании
холодом//Советская медицина. 1984. № 1. С. 92-94.
Огородникова Л. С, Гайрабедьянц Н. Г., Ратнер О. Я., Чирвина Е. Д. и
др. Морфологические кри-терии регрессии рака легкого под влиянием
магнитотерапии//Во-просы онкологии. 1980. № 1. С. 28—35.
Огородникова Л. С, Гаркав и Л. X., Андреева Л. А. Изменения в коре
надпочечников при ком-плексной химиолучевой терапии (оливомицин и
облучение) и неспецифической терапии опухолей в
эксперименте//Функционалыюе состояние желез внутренней секреции при
опухолевом процессе. Симпозиум. Тезисы докл. Ростов н/Д, 1973. С.
191-197.
Павлов И. П. Экспериментальная психология и психопатология на
животных// 20-летний опыт объективного изучения ВНД (поведения)
животных. М., 1938. С.
20-36.
:'
Павлов И. П. Павловские клинические среды. М.; Д., 1954. Т. 1. 79 с.
Панин Л. Е., Соколов В. П. Психосоматические взаимоотношения при
хроническом эмоциональ-ном напряжении. Новосибирск, 1981. 177 с.
Пашинский Г. В., Яременко К. В. Проблемы онкологической фармакотерапии. Томск, 1983. 201 с.
Петков В. Лекарство, организм, фармакологический эффект. София, 1974.
Питтендрих К. Циркадные системы: захватывание//Биологические ритмы. М.,
1984. Т. 1. С. 22 — 53. Цит. по: Путилов А. А. Указ. соч.
Плеханов Г. Ф. О восприятии человеком неощущаемых сигналов: Автореф. дне. ... канд. биол. на-ук. Томск, 1967. 22 с.
Плеханов Г. Ф. Дестабилизация неравновесных процессов как основа общего
механизма биологи-ческого действия магнитных полей//Реакции
биологических систем на магнитные поля. С. 59—80.
Пресман А. С. Электромагнитные поля и живая природа. М., 1968. 193 с.
Пригожий И. Введение в термодинамику необратимых процессов. М.; Л., 1960.
Путилов А. А. Указ. соч.
Разин М. А. Состояние иеспецифически повышенной сопротивляемости при
действии некоторых фармакологических средств//Неспецифическая
лекарственная профилактика и терапия рака. Л., 1966. С. 21—56.
Романов А. И., Стареева Л. В., Васильева В. В., Кайдаш А. Г. и др.
Исследование адаптационных реакций организма при хроническом
обструктивном бронхите как прогностический тест//Современные методы
диагностики и лечения заболеваний органов дыхания. М., 1983. С.
59—64.
,,
Романовский Ю. М., Степанова И. В., Чернавский Д. С. Что такое
математическая биофизика? (Кинетические модели в
биофизике). М., 1971. 136 с.
Рыбалко В. В., Браженко Н. А., Хавинсон В. X. Влияние тималина на
неспецифическую рози-стентность больных
туберкулезом органов дыхания//Военно-ме-
216
дицинский журнал. 1986. № 5. С. 34—36.
Самохвалова Л. И., Дерибас В. И., Белокопытов В. И. Изучение
возможности определения адапта-ционных реакций организма по измерению
потенциала акупунктурных точек//Гомеостатика жи-вых и технических
систем. С. 110 — 112.
Самунджан Е. М. Кора надпочечников и опухолевый процесс. Киев, 1973. 202 с.
Сапов И. А., Новиков В. С. Неспецифические механизмы адаптации человека. Л., 1984. 145 с.
Сауткин М. Ф., Поляков А. П., Попова М. В. Изменение белой крови под
влиянием физических нагрузок различного характера у студенток с
некоторыми забо-леваниями//Двигательная актив-ность и физическое
воспитание учащейся молодежи. Рязань, 1984. Т. 81. С. 3—5.
Селье Г. Очерки об адаптационном синдроме. М., 1960. 254 с.
Селье Г. Профилактика некрозов
сердца химическими средствами. М.,
1961.
Селье Г. На уровне целого организма. М., 1972.
Селье Г. Стресс без дистресса. М., 1979. 123 с.
Селье Г. От мечты к открытию. М., 1987. 367 с.
Серегин Ю. В. Исследование биологического действия магнитного поля на
структуры головного мозга больных с грубой органической
патологией//Приме-непие лазеров и магнитов в биологии и медицине.
Тезисы докл. С. 107.
Серег.ин [О. В. Расчет параметров магнитного поля и электрических токов
для формирования за-данного уровня неспецифическнх адаптационных
реакций организма//Там же. С. 108.
Серегин Ю, В. Разработка методов и устройства
для восстановления движении конечностей у
детей с церебральным
параличом: Автореф. дис
... канд техн наук. М., 1986. 16 с.
Серегин Ю. В., Лозин Б. А. Способ расчета параметров электрического
тока и магнитного поля но данным формулы белой крови для лечения
больных с двигательными нарушениями и нарушения-ми центрального
генеза//Магпитные поля в биологии, медицине и сельском хозяйстве.
Тезисы докл. II областной научно-практической конференции. С.
125—126.
Сетров М. И. Информационные процессы в биологических системах: Методологический очерк. Л., 1975. 155 с.
Сидякин В. Г. Влияние глобальных экологических факторов на нервную систему. Киев, 1986. 159 с.
Сидякин В. Г., Темурьянц Н. А., Евстафьева Е. В. Некоторые
биохимические и морфологические изменения крови крыс при воздействии
переменного магнитного поля инфранизкой часто-ты//Космическая биология
и авиакосмическая медицина. 1986. № 5. С. 90-91.
Симонов П. В. Три фазы в реакции организма на возрастающий стимул. М., 1962. 244 с.
Синтез фосфоенолиирувата и реакции дыхательной цепи митохондрий в
печени животных при физиологических адаптационных реакциях/Косенко Е.
В., Каминский Ю. Г., Кондрашова М. Д., Маевский Е. И. и др.//Регуляция
энергетического обмена и
физиологическое состояние
орга-низма. М., 1978.
С. 166 — 170.
Смирнов Г. В. Адаптационные реакции при восстановлении сексуальной
активности больных, пе-ренесших инфаркт миокарда//Диагностика и лечение
сексуальных расстройств. Харьков, 1983. С. 76.
Соколова Г. Б., Царькова М. Ю. Значение определения общих
неспецифических адаптационных реакций организма в клинике внутренних
болезней//Есте-ственные науки — здравоохранению. Тезисы докл.
Пермь, 1987. С. 37—38.
Сороко С. И. Нейрофизиологические механизмы индивидуальной адаптации человека в Антарк-тиде. Л:, 1984. 152 с.
Сперанский А. Д. Элементы построения теории медицины. М., 1935.
Судаков /Г. В. Системные механизмы эмоционального стресса. М., 1981. 231 с.
Тарасьянц Ф. М. Защитное влияние адаптационных реакций тренировки и
активации, вызываемых магнитным полем, при химиотерапии
опухолей//Примене-
217
ние лазеров и магнитов в биологии и медицине. Тезисы докл. С. 118.
Терапевтическое действие янтарной кислоты. Пущино,, 1976. С. 215—218.
Терещенко И. П. Резистентность организма и злокачественный рост//Тезисы
докл. II Всесоюз. съезда патофизиологов МЗ СССР. Ташкент, 1976. Т. 2.
С. 180— 190.
Тигранян Р. А. Стресс и его значение для организма. М., 1988. 175 с.
Туракулов Л. X. Щитовидная железа//Физиология эндокринной системы. Л., 1976. С. 135-191.
Удинцев Н. В., Мороз В. В. О механизме реакции
гипофизарно-надпочечни-ковой системы на стрессовое действие переменного
магнитного поля//Патологиче-екая физиология. 1976. № 6. С. 72—74.
Уколова М. А. О возможности тренировки нервных регуляций как способе
воздействия на опухо-левый процесс//Материалы конференции по
опосредованному воздействию на опухолевый про-цесс. С. 87—88.
Уколова М. А. Роль нейро-эндокринных нарушений в патогенезе опухолей яичников. М., 1972. 246 с.
Уколова М. А., Бордюшков Ю. Н., Гаркави Л. X., Гончарова В. К., Чаусов
В. И. Влияние раздраже-ний гипоталамуса на опухолевый процесс//Труды
VIII Международного противоракового кон-гресса. М.; Л., 1963. Т. 3. С.
488—490.
Уколова М. А., Веревкина Г. А. Различный механизм действия на организм
умеренных и больших доз тиоТЭФа//Вопросы клинической онкологии и
нейро-эндокринных нарушений при злокачест-венных новообразованиях. Вып.
2. С. 286 — 289.
Уколова М. А., Гаркави Л. X., Квакина Е. Б. Об изменении
взаимоотношений свертывающей и про-тивосвертывающей системы крови при
общей неспецифической адаптационной «реакции
трени-ровки»//Система свертывания крови и фиб-ринолиз. III
Всесоюз. конференция по коагулологии. Львов; Киев, 1969. С. 159—
160.
Уколова М. А., Гаркави Л. X., Квакина Е. В. О роли гипоталамуса в
приспособительных реакциях организма//Х1 съезд Всесоюз.
физиологического общества им. И. П. Павлова. М.; Л., 1970. Т. 2. С. 491.
Уколова U. А., Гаркави Л. X., Квакина Е. Б. К теоретическому
обоснованию неспецифической те-рапии опухолей//В опросы клинической
онкологии и нейроэн-докринных нарушений при злокаче-ственных
новообразованиях. Вып. 2. С. 374— 377.
Уколова М. А., Гаркави Л. X., Квакина Е. Б. Неспецифическое повышение
функциональной актив-ности эндокринных желез при химио- и лучевой
терапии опухолей//Функциональное состояние желез внутренней секреции
при опухолевом процессе. Симпозиум. Тезисы докл. Ростов н/Д, 1973. С.
162—165.
Уколова М. А., Квакина Е. Б. О влиянии магнитных полей на рост опухоли// Вопросы онкологии. 1970. № 2. С. 88—91.
Уколова М. А., Квакина Е. Б. Влияние магнитного поля на
экспериментальные опухоли (прямое и через нервную систему)//Влияние
магнитных полей на биологические объекты. М., 1971. С. 147 — 164.
Уколова М. А., Квакина Е. Б., Марьяновская Г. Я. О противоопухолевом
влиянии магнитного по-ля//Материалы II Всесоюз. совещания по изучению
влияния магнитных полей на биологические объекты. С. 237—239.
Уколова М. А., Квакина Е. Б., Рудой Ф. М. Защитное действие МП при
введении противоопухоле-вых химиопрепаратов//Там же. С. 240—241.
Уколова М, А., Квакина Е. В., Чернявская Г. Я. Энергетический обмен
гипо-таламо-гипофизарного отдела мозга крыс при противоопухолевом
влиянии магнитного поля//Вопросы онкологии. 1969. № 12. С.
60—64. ,
Ухтомский А. А. Параметры физиологической лабильности и нелинейная
теория колеба-ний//Избранные труды. Л., 1978. С. 187 — 194.
Файдыш Е. А. Моделирование динамики мотивации при инстинктивном
поведении живот-ных//Проблсмы управления в технике, экономике,
биологии. М., 1981. С. 196-203.
218
Филаретов А. А. Нервная регуляция гипофизарно-адренокортикальной системы. Л., 1979. 144 с.
Фшшпченко Р. Е., Привалов И. А., Любарская К. И., Алексеева Г. П.
Некоторые критерии оценки состояния организма в донозологической
диагностике//Про-блемы организации массовых профи-лактических осмотров
на промышленном предприятии. Новосибирск, 1980. С. 25 — 32.
Фролъкис В. В. Старение и биологические возможности организма. М. 1975 270 с.
Фролькис В. В. Биологические предпосылки увеличения продолжительности
жизни//Геронтология и гериатрия. 1975; Биологические возможности
увеличения продолжительности жизни. С. 7 —12.
Фурдуй Ф. И. Физиологические механизмы стресса и адаптации при остром действии стресс-факторов. Кишинев, 1986. 237 с.
Хайдарлиу С. X. Функциональная биохимия адаптации. Кишинев, 1984. 270 с.
Холодов Ю. А. Влияние электромагнитных и магнитных полей на ЦНС М 1966. 284 с.
Холодов Ю. А. Реакция нервной системы на электромагнитные поля. М., 1975. 208 с.
Холодов Ю. А. Мозг в электромагнитных полях. М., 1982. 120 с.
Холодов Ю. А., Шишло М. А, Электромагнитные поля в нейрофизиологии М., 1979. 168 с.
Царегородцев И. Г., Ерохин В. Г. Диалектический материализм и теоретические основы медицины. М., 1986. 288 с.
Чеботарев Д. Ф., Коркушко О. В., Калиновская Е. Г. Влияние
двигательного режима на некоторые функции организма пожилых и старых
людей//Двигательная активность и старение. С. 213—226.
Чернух А. М. Функциональный элемент — орган — организм
(особенности адаптации, поврежде-ния и восстановления)//Механизмы
повреждения, резистентности, адаптации и компенсации. Ташкент, 1976. Т.
1. С. 13—17.
Черниговский В. Н. Интерорецепторы. М., 1960. 656 с.
Чернявская Г. Я. Об энергетическом обмене гипоталамуса при росте
опухолей и некоторых проти-воопухолевых воздействиях: Автореф. дис. ...
канд мед наук Донецк, 1971. 24 с.
Чукичев И. П. Симпатомиметические вещества белкового происхождения. М., 1958.
Шакиров А. Ш. Мумие-асиль в комплексном лечении переломов костей. Ташкент, 1976. 31 с.
Шаляпина В. Г. Участие катехоламинов мозга в регуляции
гипофизарно-адреналовой систе-мы//Гипофизарно-адреналовая система и
мозг. С. 49—66.
Шварцбурд П. Н. Биофизическая характеристика лимфоцитов на разных
стадиях развития опухо-левого процесса (модель асцитной гепатомы
Зайделя): Автореф. дис. ... канд. биол. наук. Пущино, 1980. 24 с.
Шевелев А. И., Невский М. П., Гаркави Л. X., Уколова М. А. и др. О
связи антистрессорного дейст-вия реакции активации с лечебным эффектом
при психических заболеваниях//Стресс,
адаптация и функциональные
нарушения. С. 365.
Шеннон В. Имитационное моделирование систем — искусство и наука. М , 1978.
Шепотиновский В. И., Микашинович 3. И. Метаболический ответ лейкоцитов
как показатель ин-дивидуальной реакции животного на стресс и шокогенную
трав-му//Бюллетень эксперименталь-ной биологии и медицины. М., 1980. №
10. С 418 — 420.
Шихлярова А. И. О возможности передозировки при лечебном применении
слабых магнитных по-лей//Материалы III Всесоюз. симпозиума
«Влияние магнитных полей на биологические объекты».
Калининград, 1975. С. 169—170.
Шихлярова А. И. Об адаптационно-трофическом влиянии слабых магнитных
полей на половые железы старых крыс//Применение лазеров и магнитов в
биоло-
219
гии и медицине. Тезисы докл. С. 128.
Шихлярова А. И. Адаптационно-трофическое влияние^.доалых доз
адреналина: Автороф. дне. ... канд. биол. наук. Ростов н/Д, 1985. 24 с.
Шихлярова А. И. Использование математического моделирования для
прогнозирования изменений некоторых показателей резистентности
организма при развитии опухолевого процесса//! V Всесо-юз. съезд
онкологов. Тезисы докл. Л., 1986. С. "580-581.
Шихлярова А. И. О прогностической значимости регрессионных моделей для
адаптивного управ-ления организма с помощью реакции
активации//Гомеостатика живых и технических систем. С. 47 — 48.
Шихлярова А. //., Мулатова А. К., Евстратова О. Ф., Мамцева Л. И. О
неспецифическом адапта-ционно-трофическом влиянии малых доз адреналина
на половые железы старых крыс//Сравннтельная эпидемиология и
диагностика основных локализаций рака. М., 1984. С. 22 — 27.
Шишло М. Л., Кубли С. X., Нужный В. П. Биоэнергетика и регулирующие
системы организма при действии магнитных полей//Реакции биологических
систем на магнитные поля. С. 81 —102.
Шишло М. А., Кубли С. X., Шимкевич Л. Л. Формирование адаптационных
реакций организма при действии постоянных магнитных полей//Вопросы
курортологии, физиотерапии и лечебной физи-ческой культуры.
М., 1981. № 4. С. 12—18.
Шмальгаузен И. И. Кибернетические вопросы биологии. Новосибирск, 1968. 223 с.
Шноль С. Э. Синхронные в макрообъеме колебания АМФ-азной активности в
концентрированных препаратах актомиозииа//Колебательные процессы в
биологических и химических системах. Пу-щино, 1967. С. 20.
Шноль С. Э. Макроскопические флуктуации — общее свойство
биологи.ческих и физико-химических процессов. Проблемы
«дифференциальной диагностики» макроскопических флуктуа-ции
и автоколебаний. Возможная/физиологическая роль макроскопических
флуктуа-ций//Материалы Всесоюз. симпозиума «Механизмы временной
организации клетки и их регуля-ция на различных уровнях». Пущино,
1983. С. 3.
Щепин О. П., Царегородцев И. Г., Ерохин В. Г. Медицина и общество. М., 1983. 387 с.
Эверли Дж. С, Розенфельд Р. Стресс. Природа и лечение. М., 1985. 223 с.
Эмануэль Н. М. Физико-химические аспекты процесса старения и проблема
увеличения продол-жительности жизни//9-й Международный конгресс
геронтологов. Киев, 1972. Т. 1. С. 295.
Яновская А. С, Кондратьева Л. П., Слуцкий В. Б. Исследование возрастных
изменений метаболиз-ма лимфоцитов крови//Физиологическое понятие
возрастной нормы. Тезисы Всесоюз. симпо-зиума с
участием иностранных ученых. С.
38—40.
Adolph E. F. General and specitic characteristics of physiological adaptation// Am. J. Physiol. 1956. 184. 18-28.
Aschoff J., Gereche U., Wever B. Desynchronization of human circadian
rhythm// Japanes J. of Physiol-ogy. 1967. 17. 450—457.
Bahnson С. В., Bahnson M. B. Role of the ego defences: Denial and
repression in the etiology of malig-nant neoplasm//Ann. N. V. Acad.
S,ci. 1966. 125. 827 — 845.
Balitsky K. P., Vinitsky V. B. The central nervous system and cancer//Stress and cancer. Toronto, 1981. 1—43.
Baftrusch H. I. E., Stangel W. Life stress and onset of polycythaemia
vera// Cancer detect, and prevent. 1980. 3. 2. 107 — 118.
Rammer K. Stress, spread and cancer//Stress and cancer. Toronto, 1981. 137 — 104.
Bastecky J., Tondlovd #., Boleloucky Z., Stehlik J., A'ndrysek O.
Psychosoma-licke aspekty zhoubnych nadorovych onemocneni//Cas. Lek.
ces. 1984. 123. 28.
860-864.
/
Becker B. O. The effect of magnetic
fields upon the central nervous
system//
220
Biological effects of magnetic fields. 1969. 2. 207 — 214.
Biskind G. В., Bis-kind M. S. Experimental ovarian tumors in rats//Am. .1. din. Path. 1949. 19. 501.
Coping and health/Ed.: Levine S., Ursin H. N. Y.-L., 1980.
Cox T. Stress. London and Basingtoke, 1978. 199 p.
Ehret С F., Groh К. В., Meinert J. C. Circadian dyschronism and
chronotypic ecophilia as factors in ag-ing and longevity//Aging and
Biological Rhythms/Ed.: Samis H. V., Capobianco Jr., Capobianco S. N.
Y, —L., 1978.
EUertsen В., Johnsen Т., Ursin H. Relationship between the hormonal
responses to activation and cop-ing//Psychohiologv of stress. A sludv
of coping men/Kd.: Ursin H., Baad E., Levine S. N. Y.-L.,'1978. 105-124.
Frankenhaeuser M. Experimental approaches to the study of
catecholamines and emotion//Emotions: Their
Parameters and Measurement/Ed.:
Levi L. N. Y., 1975.
Gros'sarth-Malirrk
/'
TS of cancer
and internal desea-
ses//Psychother. Psychosom. 1980. 33. 122-128.
H alb erg F., Nelson W. Chronobiologic optimization of aging//Aging and
Biological Rythms/Ed.: Samis
H. V., Capobianco Jr.,
Capobianco S. N.
Y.—L., 1978.
Keim K., Sigg E. Physiological and biochemical concomitants of
restraint stress in rats//Pharmacol. Bio-chem. Behav. 1976. 4. 3.
289—297.
Lazarus B. S. Psychological stress and coping process. N. Y., 1966.
Lazarus B. S. The stress and coping paradigm/'/Competence and coping during adulthood. Hanover, 1980.
Levi L. Stress and distress in response to psychosocial stimuli//Acta med. scand., Suppl. 1973. 528.
Levine S., Weinberg /., Ursin H. Definition of the coping process and
statement of the prob-lem//Psychobiology of stress. A study of coping
men/Ed.: Ursin H., Baade E., Levine S. N. Y.-L., 1978. 3-22.
Levine S. Cortisol changes following repeated exeriences with parachute
training// Psychobiology of stress. A study of coping men/Ed.: Ursin
H., Baade E., Levine S. N. Y.-L., 1978. 51-56.
Lorenz K. On agression. L., 1966.
Mason J. W. A re-evaluation of the concept of
«non-specificity» in stress theory// J. of Psychiatric
Re-search. 1971. 8. 323—335.
Mason J. W. Emotion as reflected in patterns of endocrine
integration//Emo-tions: Their Parameters and Measurement/Ed.: Levi L.
N. Y., 1975.
Mikulaj L., Kvetnansky R., Murgas K. Changes in adrenal response during
interminent and repeated stress/'/Rev. Czechosl. Med. 1974. 20. 3. 162
— 169.
Moruzzi G., Magoun H. W. Brain stem reticular formation and activation
of the EEG//Electroencephalography and Clinical Neurosurgery. 1949. 1.
455—473.
Mohan С., Badha E. Circadian rhythma in the central cholinergic system
in aging animals//Aging and Biological Rhythms/Ed.: Samis H. V.,
Capobianco Jr. N. Y. — L., 1978.
On the mechanism of bypothalamus influence upon the tumour
process/UkoIn-va M. A., Bordjushkov J. /V., Garkavi L. H., Goncharova
V. A"., Kvakina E. li./f Acta Unio Intern. Contra Cancrum. 1964. 20.
1604 — 1607.
Persinger M. A. Effects of magnetic fields on animal behavior//Progress in biome-teorology. Amsterdam, 1976. 177 p.
Pittendrigh С S., Daan S. Circadian oscillations in rodents: A
systematic increase of their frequency with age//Science. 1974. 186.
548 — 550.
Psychobiology of stress. A study of Coping Men/Ed.: Ursin //., Baade E., Levine S. N. Y.-L., "1978. 228 p.
Selye H. Thymus and adrenals in the response of the organism to
inguries and intoxication//Brit. J. Exp. Path. 1936. 17. 234-248.
Selye H. The physiology and pathology of exposure to stress//Acta Inc. Medical Publishers. .Montreal, 1950.
Selye H. The stress of life. N. Y., 1956.
221
Selye H. The evolution of the stress concept//American Scientist. 1973. 62. 642-649.
Selye H. Stress in the health and desease//Reading Mass, Butterworth's. 1976.
Selye H. Correlation stress and cancer//Amer. J. Proctol. 1979. 30. 4.
18—20,
25-28.
Selye H. Stress, cancer and the mind//Cancer, stress and death. N. Y.—L., 1981. 11-21.
Selye H., Horava A. Third annual report on stress//Acta Inc. Medical Publishers.
Montreal, 1953.
Selye H., Tuchweber B. Stress in relation to aging and
desease//Hypothalamus, pituitary and aging. Springfield, 1976. 191
— 197.
Temoshok L. Emotion, adaptation and desease: a multidimensional
theory//Emo-tions in Health and Ill-ness: theoretical and research
foundations. N. Y., 1983. 207 — 233.
Vkolova M. A., Bordjushkov J. N., Garkavi L.Н , Goncharova V. K.,
Kvakina E. B. The effect of direct hypothalamus stimulation on the
tumor process//Acta Unio International Contra Cancrum. 1964. 20.
1604-1607.
Ursin H. Activation, coping and psychosomatics//Psychobiology of
stress. A study of coping men/Ed.:
Ursin Н., Baade E.,
Levine S. N.. Y.-L.,
1978. 201-228.
Vigas M. Problems of definition of stress stimulus and specificity of
stress respon-se//Stress: The role of catecholamines and other
neurotransmitters. N. Y., 1984. 1. 27-35.
Wever R. The meaning of circadian rhythmicity with regard to
aging//Verhand-lungen der Deutschen Ge-sellschaft fur Pathologie. 1975.
59. 160—180.
224
ОГЛАВЛЕНИЕ
Введение ........................ 3
Адаптационные реакции организма
(Л. X. Гаркави. Е.
Б. Квакина. М. А.
Уколов»)
.... 5
Общая неспецифическая
реакция организма
на сильные раздражители —
стресс (реак-ция
напряжении)
..................... 10
О возможности существования
других (кроме стресса)
неспецифических адаптационных
реакций
организма)....................
14
Реакция тренировки — общая
неспецифическая адаптационная
реакция на слабые раздражите ли
(Л. X. Гаркави, Е.
Б. Квакина, М. А.
Уколова).............. 17
Резистентиость организма при
развитии реакции тренировки
......... 32
Реакция активации — общая
неспецифическая адаптационная
реакция на раздражи-тели
«средней»
силы (Л. X. Гаркави)..................... 36
Резистентиость организма при развитии реакции активации.......... 51
О принципе периодичности
в развитии адаптационных
реакций и ареактнвностн
(Л. X. Гаркви.Е. Б.
Квакина)....................... 64
Периодическая закономерность
развития общих адаптационных
реакций...... 64
Периодическая закономерность развития местных
адаптационных реакций...... 82
Сочетание неспецифических и
специфических реакций
........... 86
Синхронизация, резонансные явления,
энтропия, негэнтропия в
периодической системе
адаптационных
реакций.................... 88
Система ареактнвности в
поддержании
гомеостаза............ 93
Представление о норме, предболезни, болезни, резервах
и старении организма в связи с
различными типами адаптационных
реакций (Л. X. Гаркави,
Е. Б. Квакина, М. А.
Уколова) .... 101
Тип адаптационных
реакций и
норма............... 101
Тип адаптационных реакций
и резервы
организма............ 102
Тип адаптационных реакций и старение............... 106
Представление о норме, предболезни,
болезни, и резервах и
старении организма в связи с
многоуровневостью развития адаптационных
реакций и ареактивности (Л. X.
Гаркави, Е. Б. Квакина) . 111
Норма, предболезнь и болезнь
с учетом реакций разных
уровней реактивности и со-стояний
ареактнвности
................ 111
Резервы организма с учетом реакций разных
уровнен реактивности и ареактнвности … 115
Старение с учетом реакций разных уровней реактивности и ареактнвности ...... 116
Принципы применения реакций активации и
тренировки для профилактики и
лечения (Л. X. Гаркави,
Е. Б. Квакина, М.
А. Уколова)................. 124
Принципы использования реакций актпвации и тренировки
разных уровней реактивно-сти для оздоровления,
профилактики и лечения (Л. X. Гаркави, Е.
Б. Квакина)..........133
Повышение сопротивляемости
организма с помощью
адаптационных реакций тренировки и
активации на разных
уровнях реактивности
организма (активационная
терапия) на основе обратной связи с
организмом .................. 135
О программированных режимах
и их месте в
активационных оздоровлении, профилак-тике
и лечении....................... 143
Применение активационной терапии для
профилактики и
оздоровлении
...... 150
О принципе диагностического лечения
с целью выявления групп
риска и лечения
забо-левания на
начальных стадиях..................
156
Применение активационной терапии на разных этапах лечения больных с опухолями …. 158
Математические модели................... 160
Дальнейшее развитие и применение
теории адаптационных реакций разных
уровнен реак-тивности в
отдельных областях
медицины............... 161
К теории адаптационных реакций
(Л. X. Гаркави, Е. Б.
Квакина, М. А. Уколова).... 169
К теории многоуровневой регуляции
гомеостаза организма с
помощью периодической системы
неспецифических адаптационных реакций и ареактнвностн (Л. X. Гаркави,
Е. Б. Квакина) .
.
.
176
Заключение....................... 194
Литература........................ 203
|
|
|
|

